

Early elevated IFNα is a key mediator of HIV pathogenesis
- PMID: 38504106
- PMCID: PMC10951235
- DOI: 10.1038/s43856-024-00454-6
Early elevated IFNα is a key mediator of HIV pathogenesis
Abstract
Background: A complete understanding of the different steps of HIV replication and an effective drug combination have led to modern antiretroviral regimens that block HIV replication for decades, but these therapies are not curative and must be taken for life. "Elite controllers" (ECs) is a term for the 0.5% of HIV-infected persons requiring no antiretroviral therapy, whose status may point the way toward a functional HIV cure. Defining the mechanisms of this control may be key to understanding how to replicate this functional cure in others.
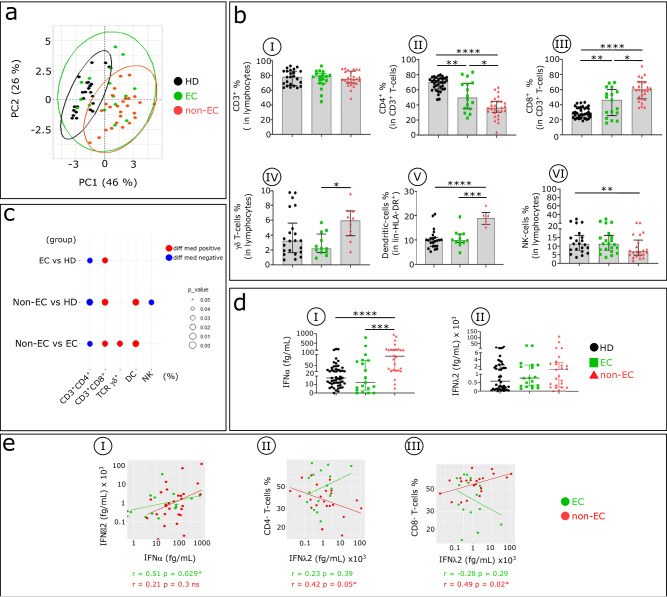
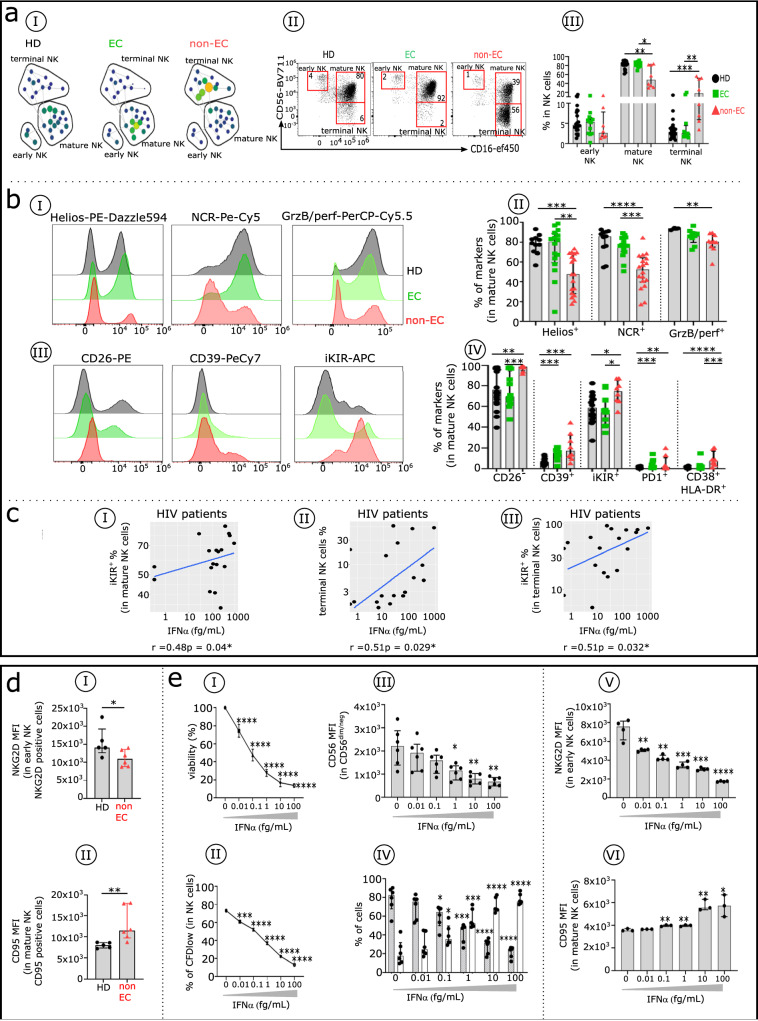
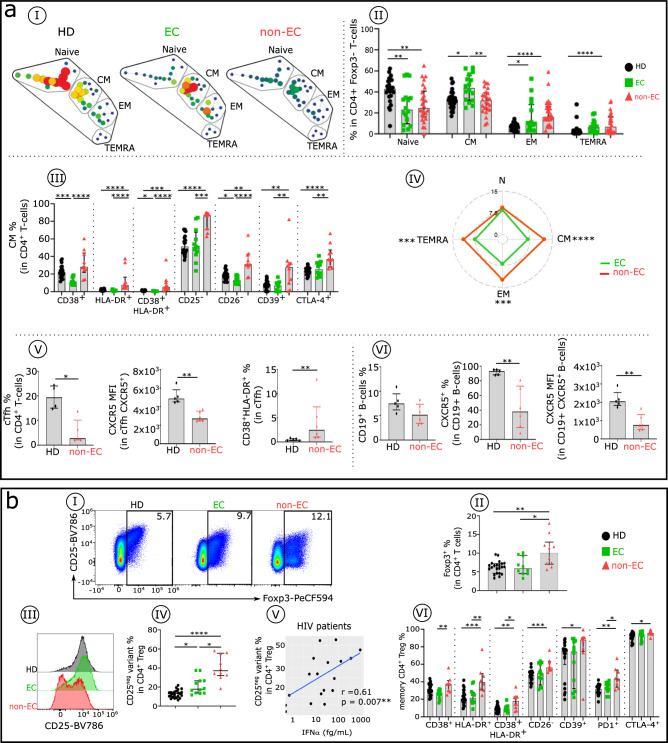
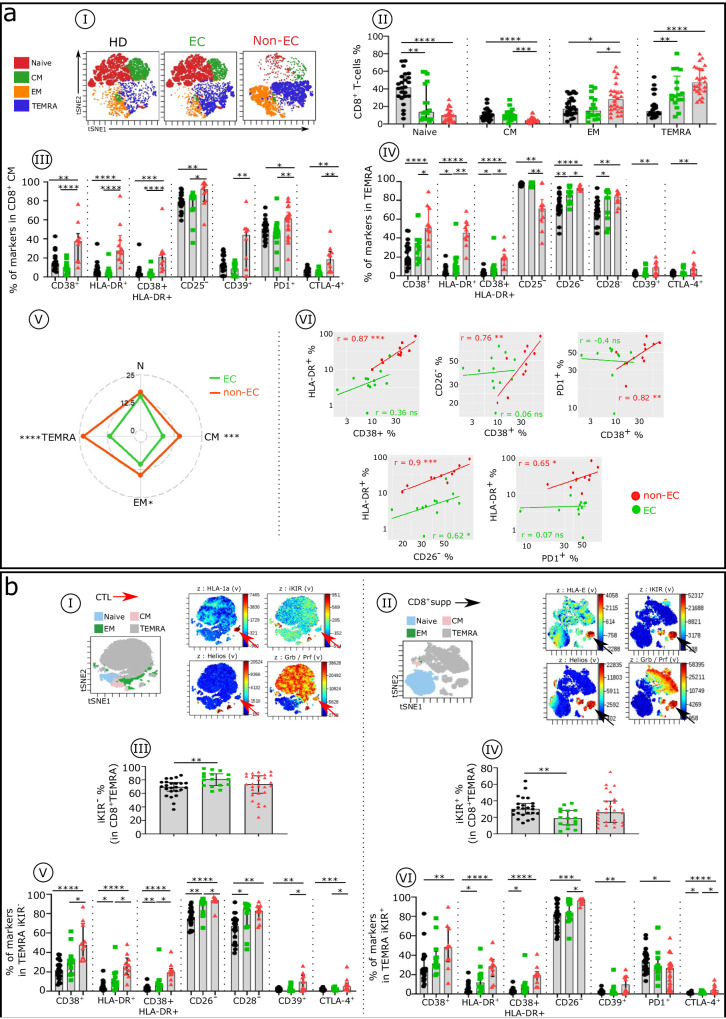
Methods: In ECs and untreated non-EC patients, we compared IFNα serum concentration, distribution of immune cell subsets, and frequency of cell markers associated with immune dysfunction. We also investigated the effect of an elevated dose of IFNα on distinct subsets within dendritic cells, natural killer cells, and CD4+ and CD8 + T cells.
Results: Serum IFNα was undetectable in ECs, but all immune cell subsets from untreated non-EC patients were structurally and functionally impaired. We also show that the altered phenotype and function of these cell subsets in non-EC patients can be recapitulated when cells are stimulated in vitro with high-dose IFNα.
Conclusions: Elevated IFNα is a key mediator of HIV pathogenesis.
Plain language summary
Currently, HIV infection is not curable, but infected individuals can manage their condition by taking daily doses of antiretroviral therapy. Some individuals, known as elite controllers (ECs), control their infection without antiretroviral treatment, and studying how their immune system responds to HIV exposure could lead to a potential cure for others. Here, we compare immune cell responses between ECs and untreated non-ECs. We find that IFNα, a small protein with an important role in controlling white blood cell activity, is produced in excess in immune cells from non-ECs compared with ECs during early infection. This insight provides an important clue for the future development of a targeted cure for HIV.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Update of
-
Early Elevated IFNα Identified as the Key Mediator of HIV Pathogenesis and its low level a Hallmark of Elite Controllers.Res Sq [Preprint]. 2023 May 11:rs.3.rs-2813601. doi: 10.21203/rs.3.rs-2813601/v1. Res Sq. 2023. Update in: Commun Med (Lond). 2024 Mar 19;4(1):53. doi: 10.1038/s43856-024-00454-6. PMID: 37215045 Free PMC article. Updated. Preprint.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
