

Gastrulation-stage gene expression in Nipbl+/- mouse embryos foreshadows the development of syndromic birth defects
- PMID: 38507484
- PMCID: PMC10954218
- DOI: 10.1126/sciadv.adl4239
Gastrulation-stage gene expression in Nipbl+/- mouse embryos foreshadows the development of syndromic birth defects
Abstract
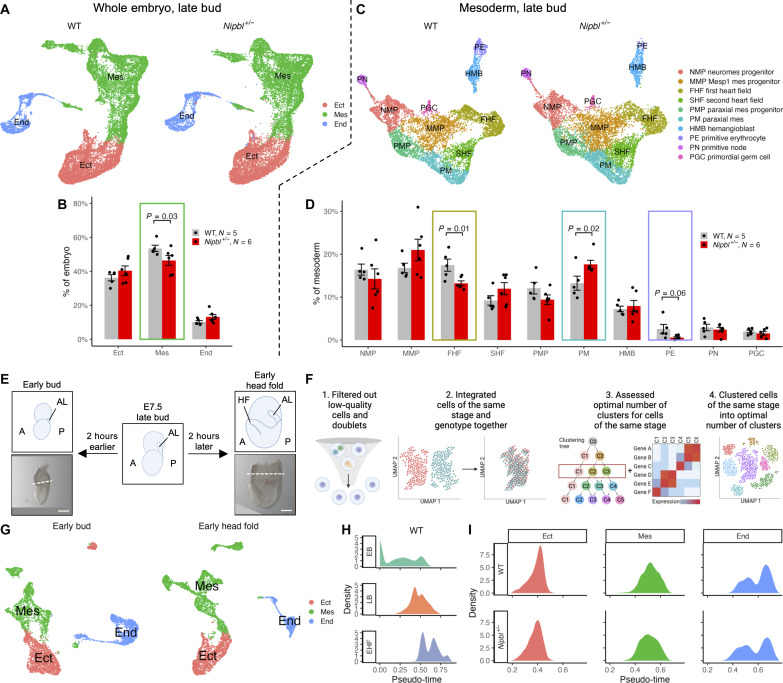
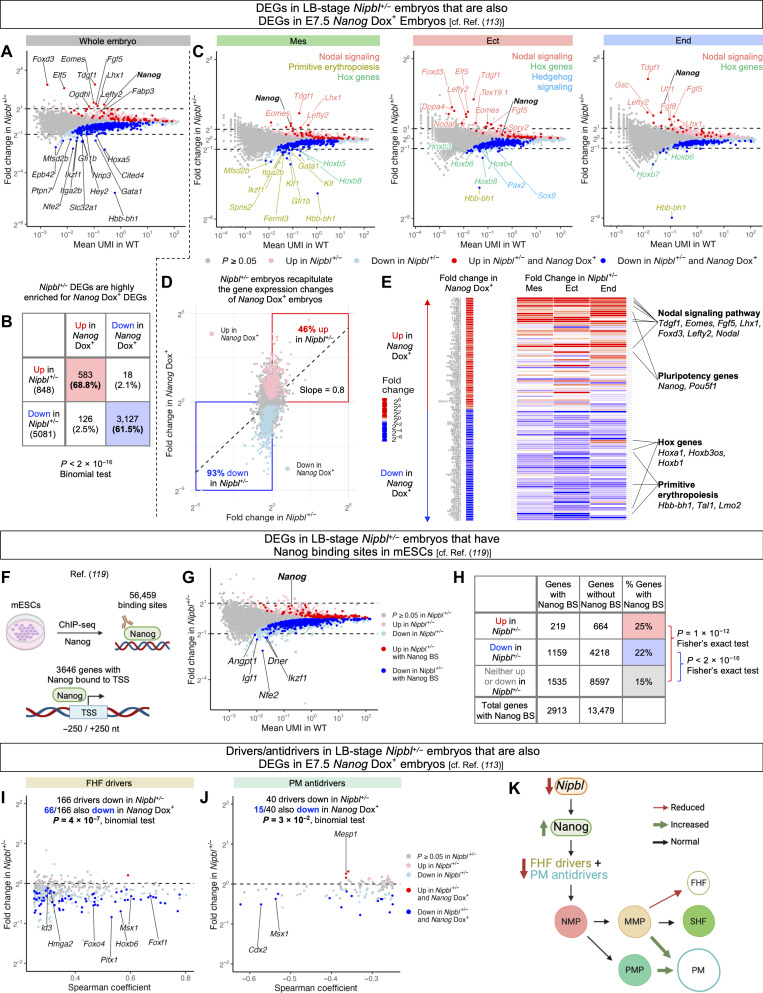
In animal models, Nipbl deficiency phenocopies gene expression changes and birth defects seen in Cornelia de Lange syndrome, the most common cause of which is Nipbl haploinsufficiency. Previous studies in Nipbl+/- mice suggested that heart development is abnormal as soon as cardiogenic tissue is formed. To investigate this, we performed single-cell RNA sequencing on wild-type and Nipbl+/- mouse embryos at gastrulation and early cardiac crescent stages. Nipbl+/- embryos had fewer mesoderm cells than wild-type and altered proportions of mesodermal cell subpopulations. These findings were associated with underexpression of genes implicated in driving specific mesodermal lineages. In addition, Nanog was found to be overexpressed in all germ layers, and many gene expression changes observed in Nipbl+/- embryos could be attributed to Nanog overexpression. These findings establish a link between Nipbl deficiency, Nanog overexpression, and gene expression dysregulation/lineage misallocation, which ultimately manifest as birth defects in Nipbl+/- animals and Cornelia de Lange syndrome.
Figures








Update of
-
Gastrulation-stage gene expression in Nipbl +/- mouse embryos foreshadows the development of syndromic birth defects.bioRxiv [Preprint]. 2024 Feb 13:2023.10.16.558465. doi: 10.1101/2023.10.16.558465. bioRxiv. 2024. Update in: Sci Adv. 2024 Mar 22;10(12):eadl4239. doi: 10.1126/sciadv.adl4239. PMID: 37905011 Free PMC article. Updated. Preprint.
References
-
- Centers for Disease Control and Prevention , Update on overall prevalence of major birth defects–Atlanta, Georgia, 1978–2005. MMWR Morb. Mortal. Wkly. Rep. 57, 1–5 (2008). - PubMed
-
- Mai C. T., Isenburg J. L., Canfield M. A., Meyer R. E., Correa A., Alverson C. J., Lupo P. J., Riehle-Colarusso T., Cho S. J., Aggarwal D., Kirby R. S., National Birth Defects Prevention Network , National population-based estimates for major birth defects, 2010-2014. Birth Defects Res 111, 1420–1435 (2019). - PMC - PubMed
-
- Sun R., Liu M., Lu L., Zheng Y., Zhang P., Congenital heart disease: Causes, diagnosis, symptoms, and treatments. Cell Biochem. Biophys. 72, 857–860 (2015). - PubMed
-
- Ely D. M., Driscoll A. K., Infant mortality in the United States, 2020: Data from the period linked birth/infant death file. Natl. Vital Stat. Rep. 71, 1–18 (2022). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
