

Energy balance drives diurnal and nocturnal brain transcriptome rhythms
- PMID: 38508192
- PMCID: PMC11330649
- DOI: 10.1016/j.celrep.2024.113951
Energy balance drives diurnal and nocturnal brain transcriptome rhythms
Abstract
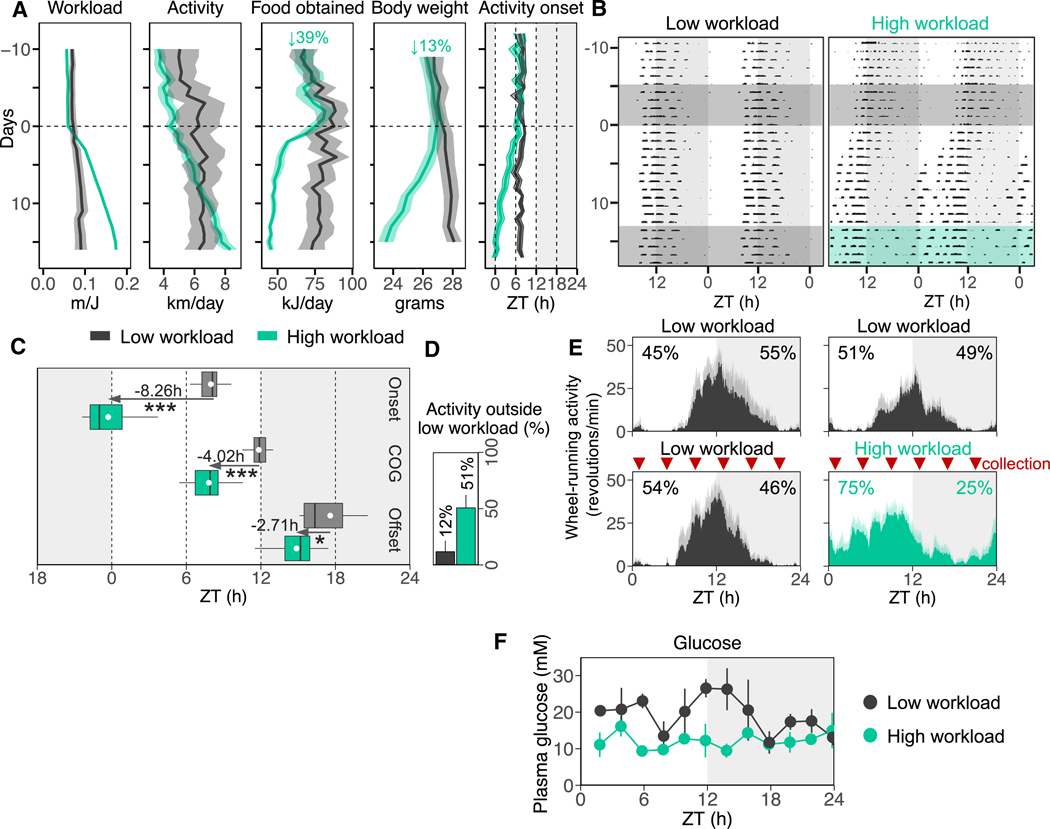
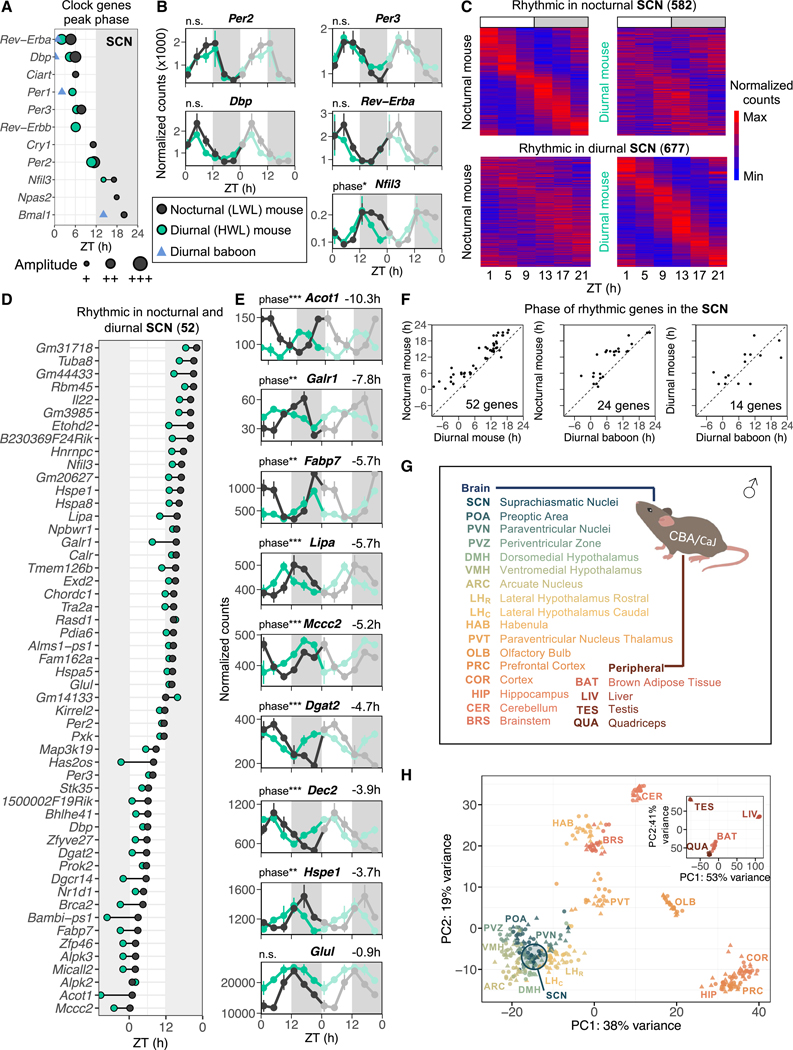
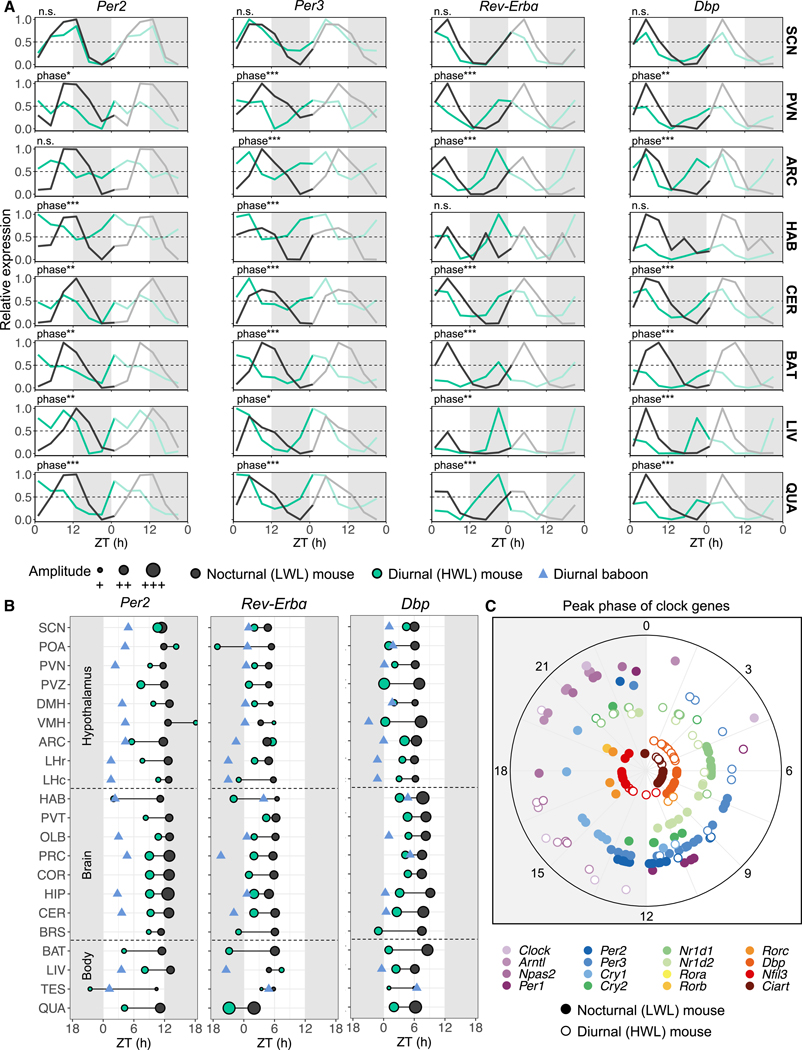
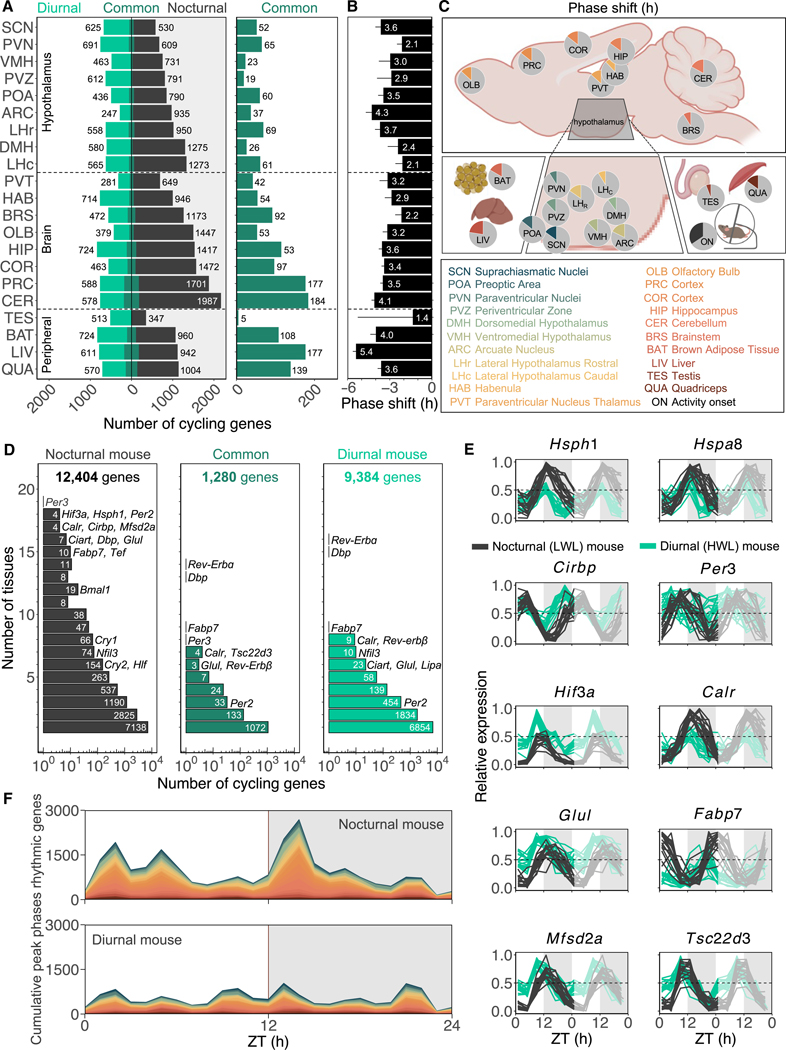
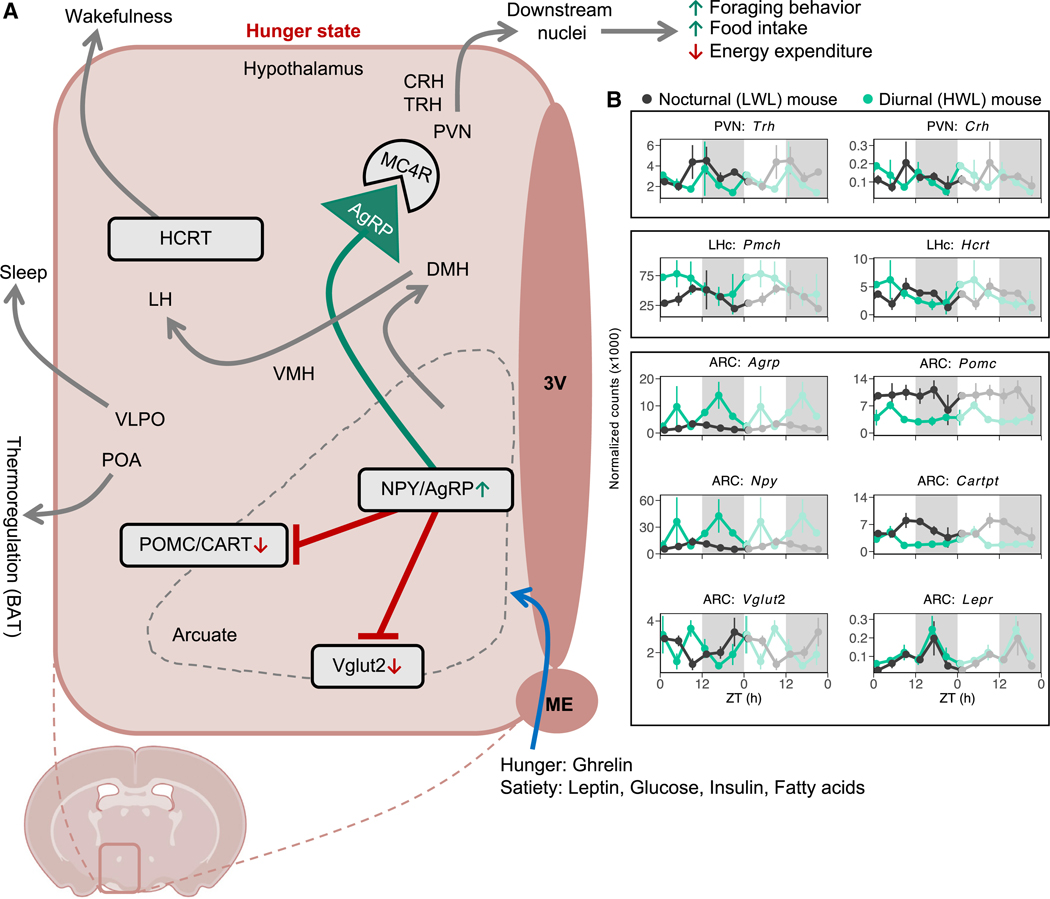
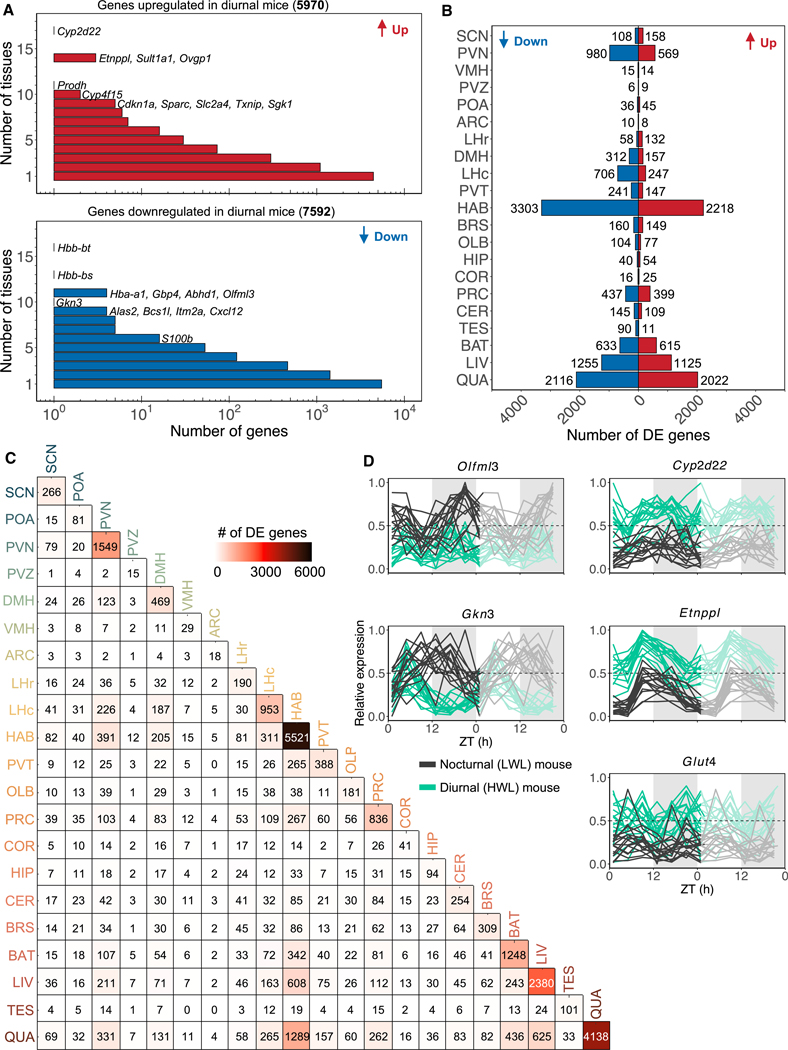
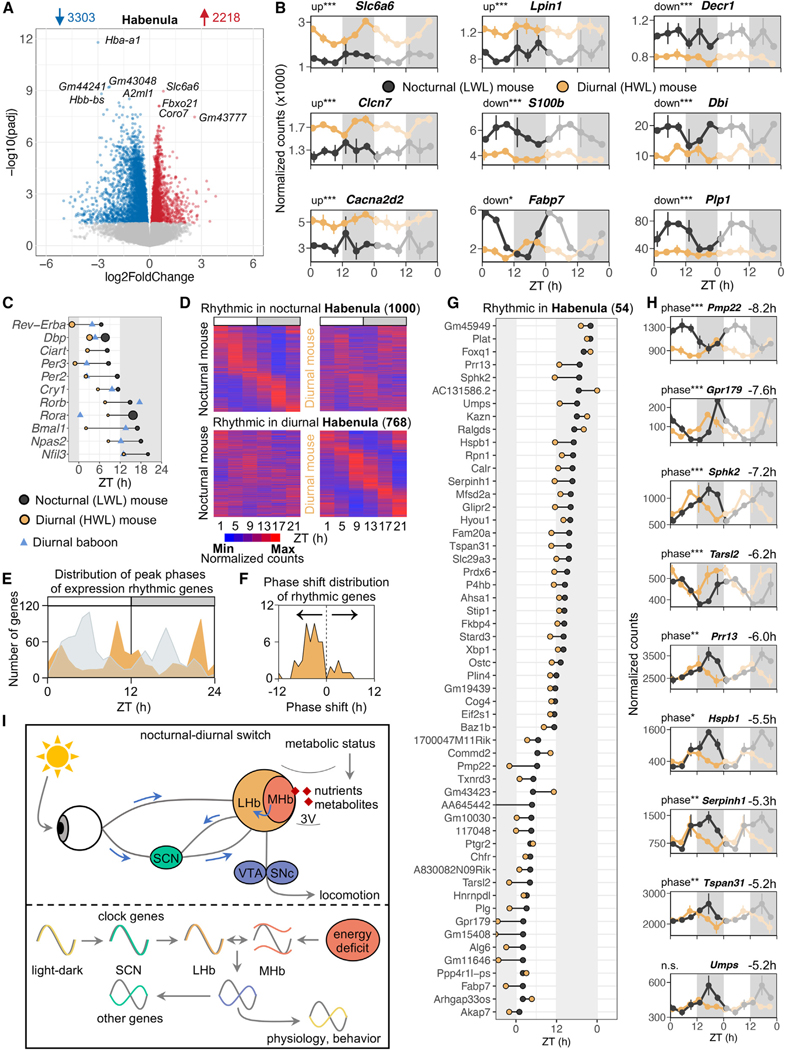
Plasticity in daily timing of activity has been observed in many species, yet the underlying mechanisms driving nocturnality and diurnality are unknown. By regulating how much wheel-running activity will be rewarded with a food pellet, we can manipulate energy balance and switch mice to be nocturnal or diurnal. Here, we present the rhythmic transcriptome of 21 tissues, including 17 brain regions, sampled every 4 h over a 24-h period from nocturnal and diurnal male CBA/CaJ mice. Rhythmic gene expression across tissues comprised different sets of genes with minimal overlap between nocturnal and diurnal mice. We show that non-clock genes in the suprachiasmatic nucleus (SCN) change, and the habenula was most affected. Our results indicate that adaptive flexibility in daily timing of behavior is supported by gene expression dynamics in many tissues and brain regions, especially in the habenula, which suggests a crucial role for the observed nocturnal-diurnal switch.
Keywords: CP: Metabolism; CP: Neuroscience; SCN; behavioral plasticity; circadian rhythms; diurnal; habenula; hypothalamus; negative energy balance; nocturnal; running wheel activity; temporal niche switching.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
