

Pregnenolone sulfate induces transcriptional and immunoregulatory effects on T cells
- PMID: 38514798
- PMCID: PMC10958000
- DOI: 10.1038/s41598-024-57327-0
Pregnenolone sulfate induces transcriptional and immunoregulatory effects on T cells
Abstract
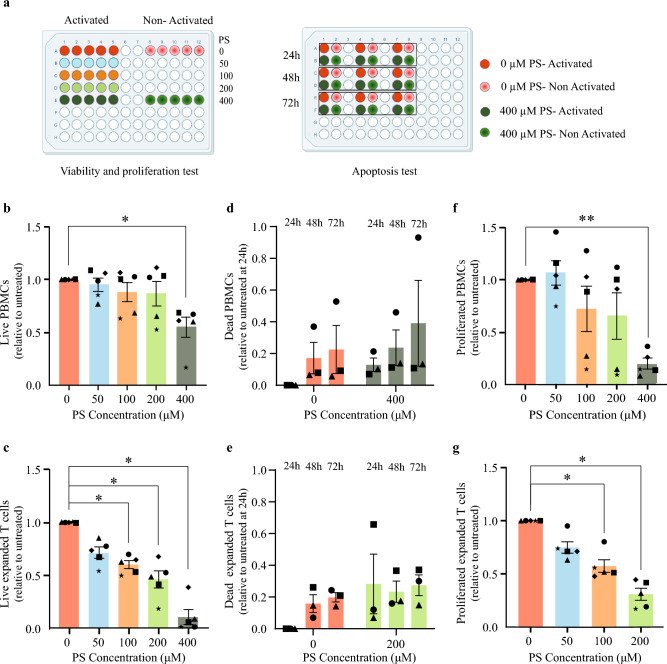
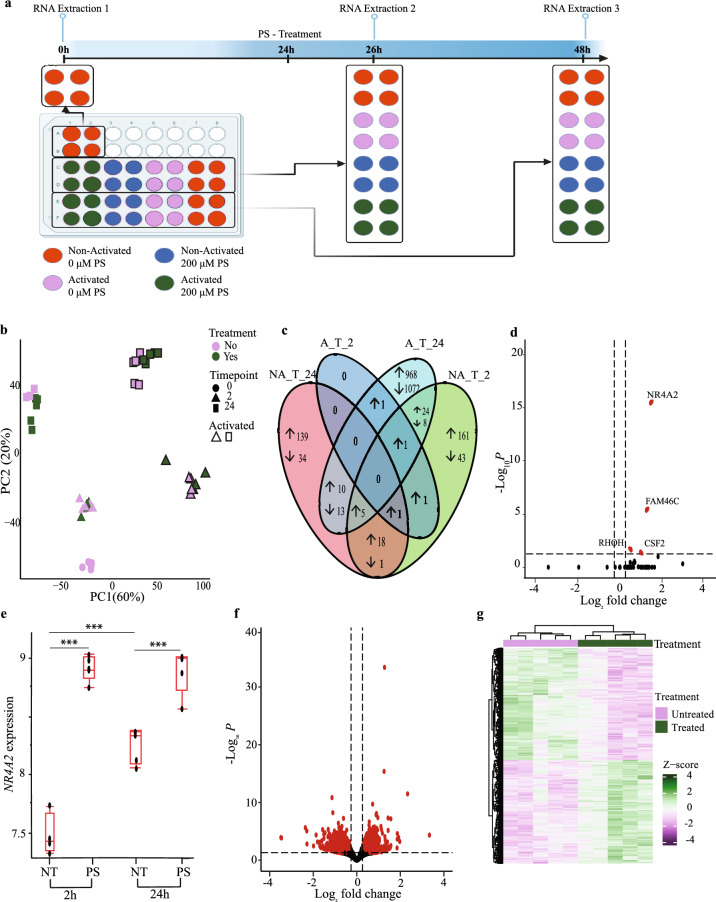
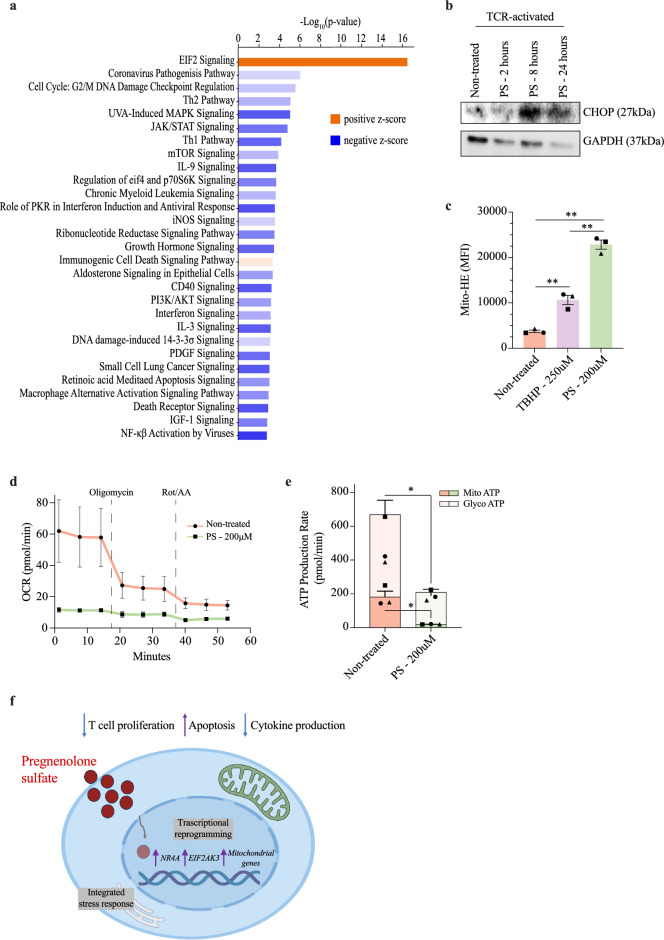
Pregnenolone sulfate is a steroid metabolite of the steroidogenesis precursor, pregnenolone, with similar functional properties, including immunosuppression. We recently reported an elevation in serum levels of pregnenolone sulfate in children with malaria, contributing to an immunosuppressed state. Yet, the molecular mechanisms in which this steroid exerts its immunoregulatory functions are lacking. In this study, we examined the effects of pregnenolone sulfate on T cell viability, proliferation and transcriptome. We observed a pregnenolone sulfate dose-dependent induction of T cell death and reduction in proliferation. RNA sequencing analysis of pregnenolone sulfate-treated T cells for 2 and 24 h revealed the downregulation of pro-inflammatory genes and the upregulation of the steroid nuclear receptor superfamily, NR4A, as early-response genes. We also report a strong activation of the integrated stress response mediated by the upregulation of EIF2AK3. These results contribute to the knowledge on transcriptional regulation driving the immunoregulatory effects of pregnenolone sulfate on T cells.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
