

Cell cycle perturbation uncouples mitotic progression and invasive behavior in a post-mitotic cell
- PMID: 38522217
- PMCID: PMC11196158
- DOI: 10.1016/j.diff.2024.100765
Cell cycle perturbation uncouples mitotic progression and invasive behavior in a post-mitotic cell
Abstract
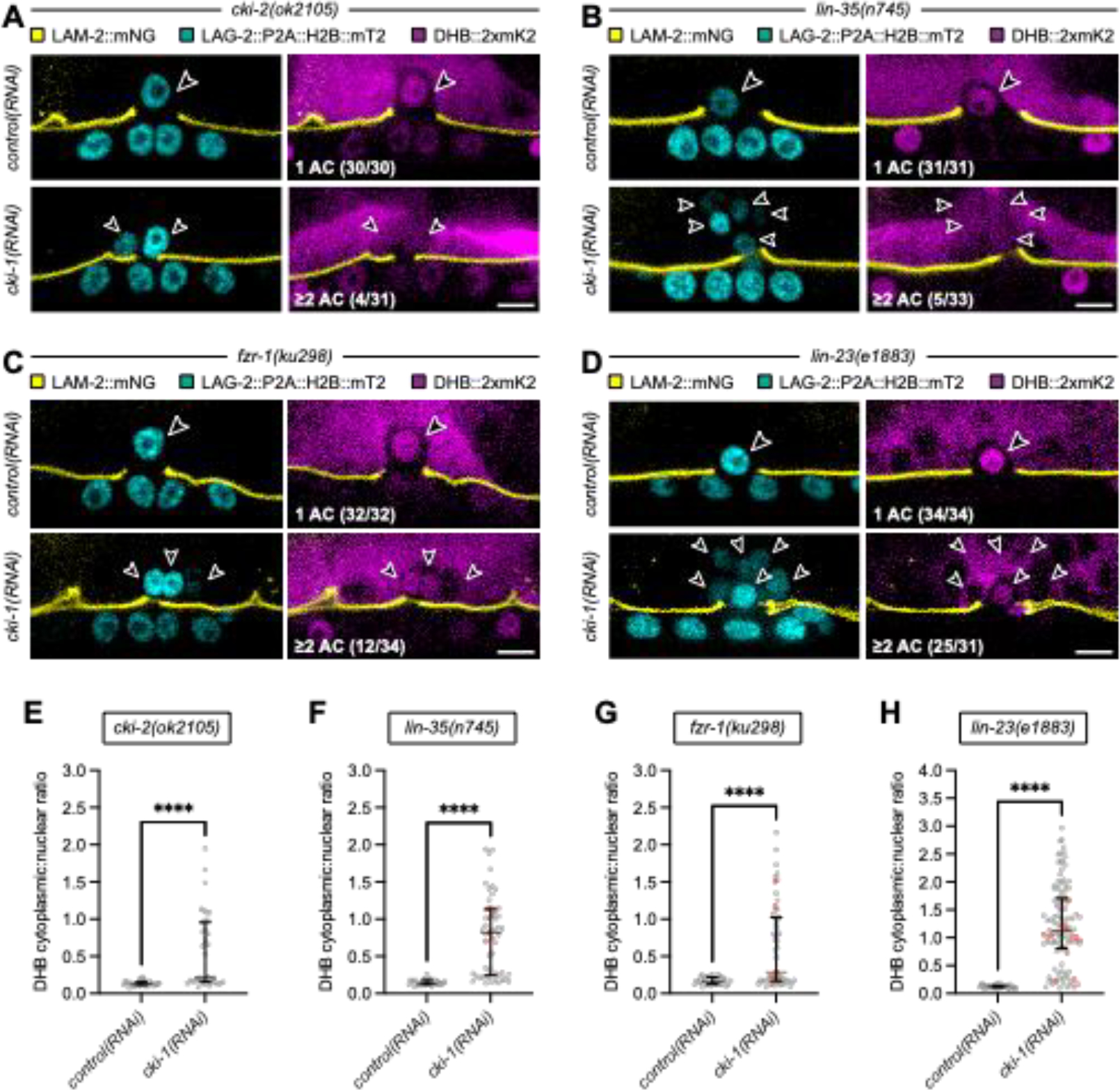
The acquisition of the post-mitotic state is crucial for the execution of many terminally differentiated cell behaviors during organismal development. However, the mechanisms that maintain the post-mitotic state in this context remain poorly understood. To gain insight into these mechanisms, we used the genetically and visually accessible model of C. elegans anchor cell (AC) invasion into the vulval epithelium. The AC is a terminally differentiated uterine cell that normally exits the cell cycle and enters a post-mitotic state before initiating contact between the uterus and vulva through a cell invasion event. Here, we set out to identify the set of negative cell cycle regulators that maintain the AC in this post-mitotic, invasive state. Our findings revealed a critical role for CKI-1 (p21CIP1/p27KIP1) in redundantly maintaining the post-mitotic state of the AC, as loss of CKI-1 in combination with other negative cell cycle regulators-including CKI-2 (p21CIP1/p27KIP1), LIN-35 (pRb/p107/p130), FZR-1 (Cdh1/Hct1), and LIN-23 (β-TrCP)-resulted in proliferating ACs. Remarkably, time-lapse imaging revealed that these ACs retain their ability to invade. Upon examination of a node in the gene regulatory network controlling AC invasion, we determined that proliferating, invasive ACs do so by maintaining aspects of pro-invasive gene expression. We therefore report that the requirement for a post-mitotic state for invasive cell behavior can be bypassed following direct cell cycle perturbation.
Keywords: C. elegans; CKI-1; Dichotomy; Invasion; Proliferation.
Copyright © 2024 International Society of Differentiation. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures




Update of
-
Cell cycle perturbation uncouples mitotic progression and invasive behavior in a post-mitotic cell.bioRxiv [Preprint]. 2024 Feb 7:2023.03.16.533034. doi: 10.1101/2023.03.16.533034. bioRxiv. 2024. Update in: Differentiation. 2024 May-Jun;137:100765. doi: 10.1016/j.diff.2024.100765. PMID: 38370624 Free PMC article. Updated. Preprint.
References
-
- Adikes RC, Kohrman AQ, Martinez MAQ, Palmisano NJ, Smith JJ, Medwig-Kinney TN, Min M, Sallee MD, Ahmed OB, Kim N, Liu S, Morabito RD, Weeks N, Zhao Q, Zhang W, Feldman JL, Barkoulas M, Pani AM, Spencer SL, Martin BL, Matus DQ, 2020. Visualizing the metazoan proliferation-quiescence decision in vivo. eLife 9, e63265. 10.7554/eLife.63265 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
