

An oocyte-specific Cas9-expressing mouse for germline CRISPR/Cas9-mediated genome editing
- PMID: 38523431
- PMCID: PMC10987075
- DOI: 10.1002/dvg.23589
An oocyte-specific Cas9-expressing mouse for germline CRISPR/Cas9-mediated genome editing
Abstract
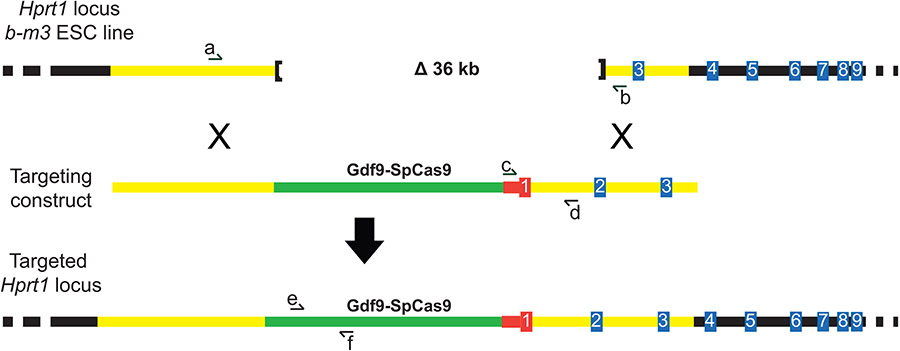
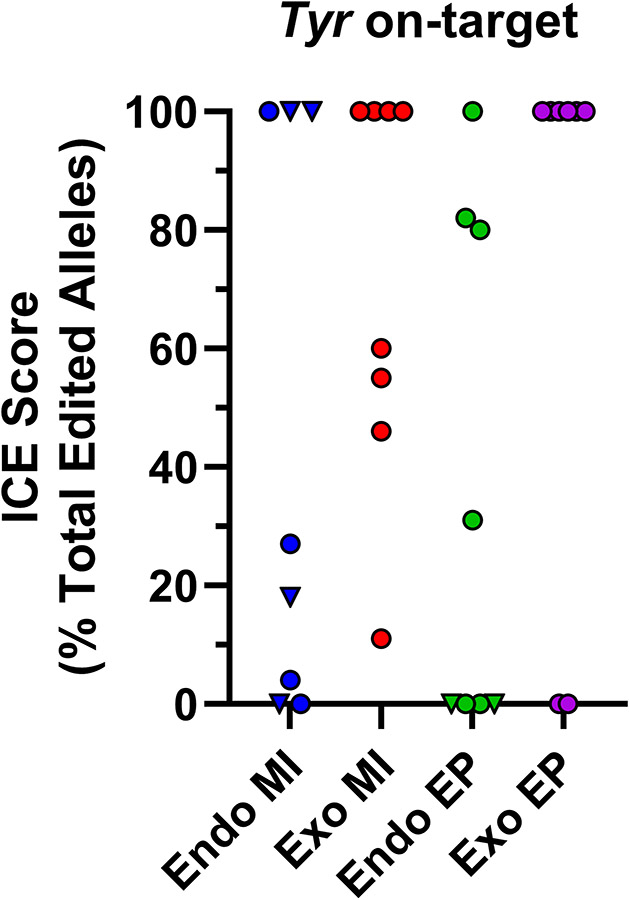
Cas9 transgenes can be employed for genome editing in mouse zygotes. However, using transgenic instead of exogenous Cas9 to produce gene-edited animals creates unique issues including ill-defined transgene integration sites, the potential for prolonged Cas9 expression in transgenic embryos, and increased genotyping burden. To overcome these issues, we generated mice harboring an oocyte-specific, Gdf9 promoter driven, Cas9 transgene (Gdf9-Cas9) targeted as a single copy into the Hprt1 locus. The X-linked Hprt1 locus was selected because it is a defined integration site that does not influence transgene expression, and breeding of transgenic males generates obligate transgenic females to serve as embryo donors. Using microinjections and electroporation to introduce sgRNAs into zygotes derived from transgenic dams, we demonstrate that Gdf9-Cas9 mediates genome editing as efficiently as exogenous Cas9 at several loci. We show that genome editing efficiency is independent of transgene inheritance, verifying that maternally derived Cas9 facilitates genome editing. We also show that paternal inheritance of Gdf9-Cas9 does not mediate genome editing, confirming that Gdf9-Cas9 is not expressed in embryos. Finally, we demonstrate that off-target mutagenesis is equally rare when using transgenic or exogenous Cas9. Together, these results show that the Gdf9-Cas9 transgene is a viable alternative to exogenous Cas9.
Keywords: CRISPR; SpCas9; genome editing; germline mutations.
© 2024 Wiley Periodicals LLC.
Conflict of interest statement
Conflict of interest: The authors have no financial or non-financial competing interests to declare.
Figures








References
-
- Alghadban S, Bouchareb A, Hinch R, Hernandez-Pliego P, Biggs D, Preece C, & Davies B (2020). Electroporation and genetic supply of Cas9 increase the generation efficiency of CRISPR/Cas9 knock-in alleles in C57BL/6J mouse zygotes. Sci Rep, 10(1), 17912. 10.1038/s41598-020-74960-7 - DOI - PMC - PubMed
-
- Bronson SK, Plaehn EG, Kluckman KD, Hagaman JR, Maeda N, & Smithies O (1996). Single-copy transgenic mice with chosen-site integration. Proc.Natl.Acad.Sci.U.S.A, 93(17), 9067–9072. http://www.ncbi.nlm.nih.gov/pubmed/8799155 (Not in File) - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
