

This is a preprint.
An injury-induced tissue niche shaped by mesenchymal plasticity coordinates the regenerative and disease response in the lung
- PMID: 38529490
- PMCID: PMC10962740
- DOI: 10.1101/2024.02.26.582147
An injury-induced tissue niche shaped by mesenchymal plasticity coordinates the regenerative and disease response in the lung
Abstract
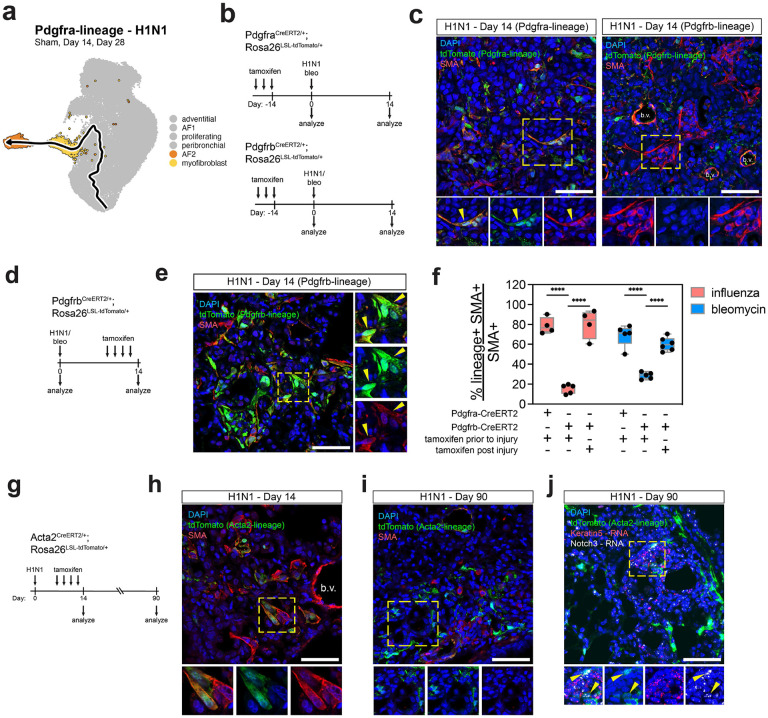
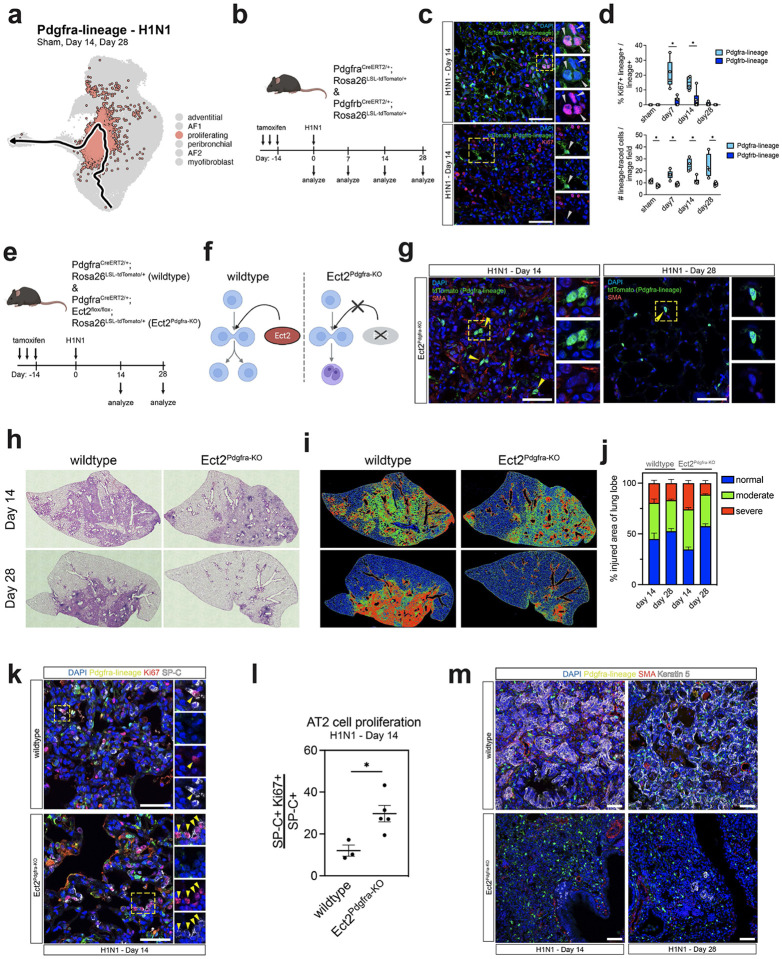
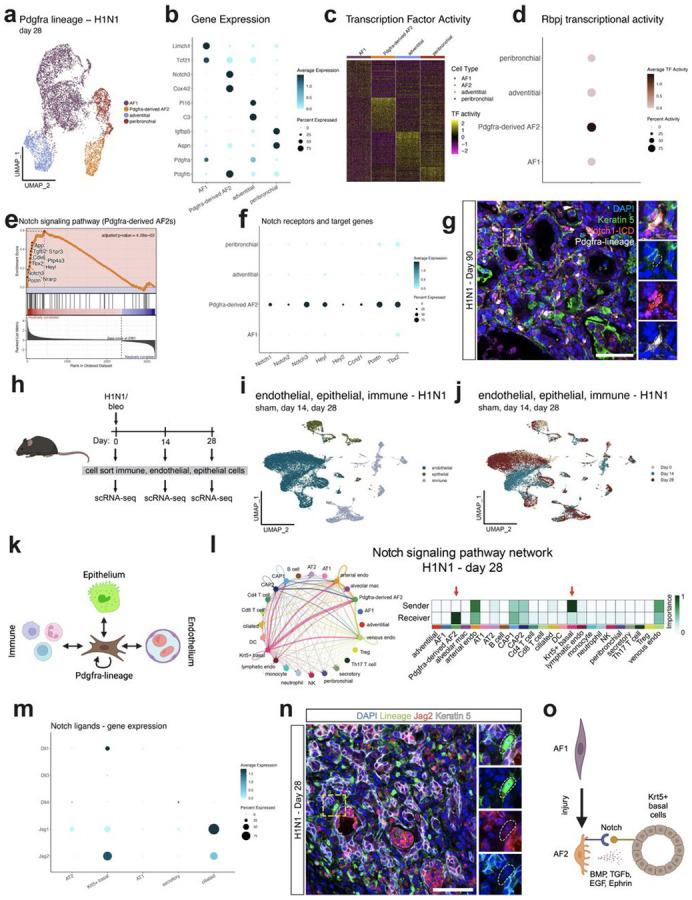
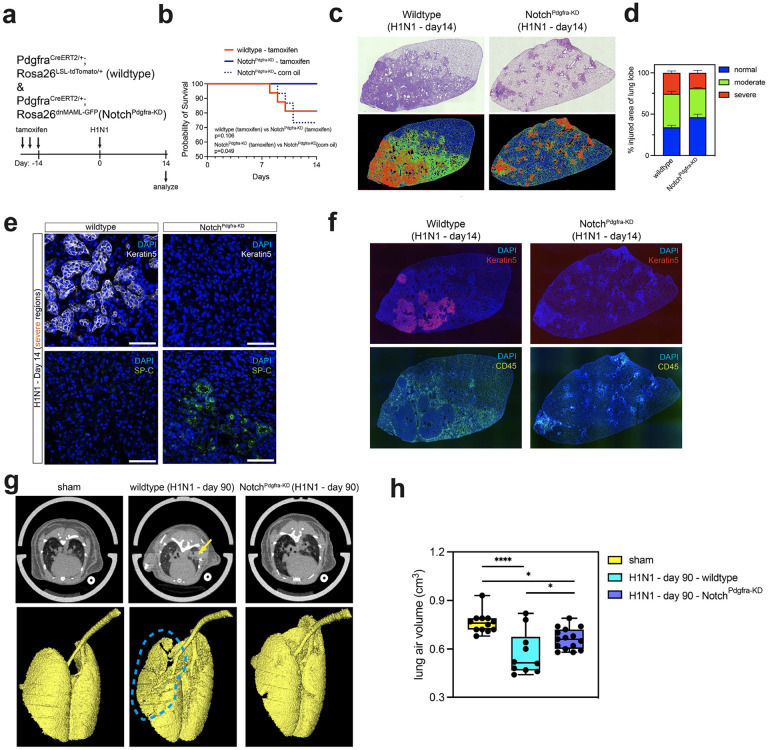
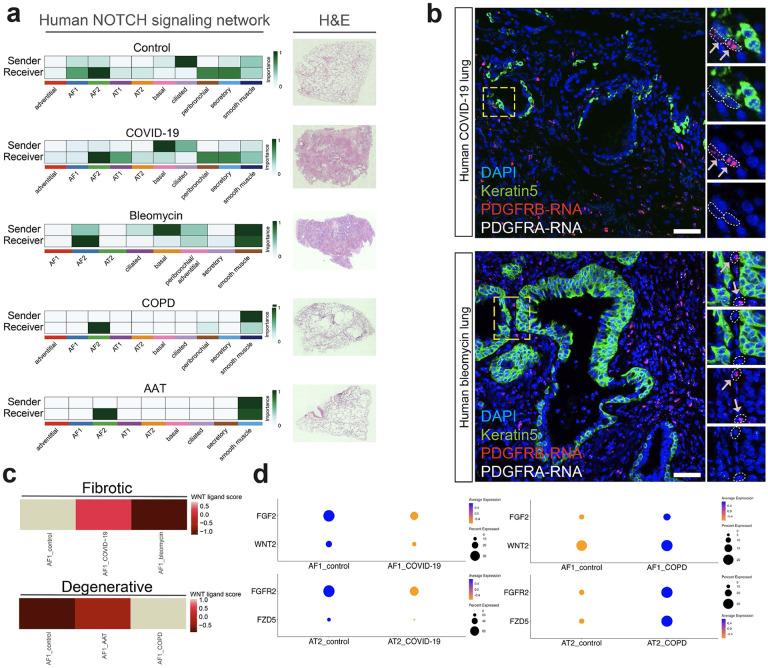
Severe lung injury causes basal stem cells to migrate and outcompete alveolar stem cells resulting in dysplastic repair and a loss of gas exchange function. This "stem cell collision" is part of a multistep process that is now revealed to generate an injury-induced tissue niche (iTCH) containing Keratin 5+ epithelial cells and plastic Pdgfra+ mesenchymal cells. Temporal and spatial single cell analysis reveals that iTCHs are governed by mesenchymal proliferation and Notch signaling, which suppresses Wnt and Fgf signaling in iTCHs. Conversely, loss of Notch in iTCHs rewires alveolar signaling patterns to promote euplastic regeneration and gas exchange. The signaling patterns of iTCHs can differentially phenotype fibrotic from degenerative human lung diseases, through apposing flows of FGF and WNT signaling. These data reveal the emergence of an injury and disease associated iTCH in the lung and the ability of using iTCH specific signaling patterns to discriminate human lung disease phenotypes.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures


References
-
- Wei X., Narasimhan H., Zhu B., Sun J., Host Recovery from Respiratory Viral Infection. Annual Review of Immunology 41, 277–300 (2023). - PubMed
-
- Boyd D. F., Allen E. K., Randolph A. G., Guo X.-Z. J., Weng Y., Sanders C. J., Bajracharya R., Lee N. K., Guy C. S., Vogel P., Guan W., Li Y., Liu X., Novak T., Newhams M. M., Fabrizio T. P., Wohlgemuth N., Mourani P. M., PALISI Pediatric Intensive Care Influenza (PICFLU) Investigators, Wight T. N., Schultz-Cherry S., Cormier S. A., Shaw-Saliba K., Pekosz A., Rothman R. E., Chen K.-F., Yang Z., Webby R. J., Zhong N., Crawford J. C., Thomas P. G., Exuberant fibroblast activity compromises lung function via ADAMTS4. Nature 587, 466–471 (2020). - PMC - PubMed
-
- Basil M. C., Katzen J., Engler A. E., Guo M., Herriges M. J., Kathiriya J. J., Windmueller R., Ysasi A. B., Zacharias W. J., Chapman H. A., Kohon D. N., Rock J. R., Snoeck H. W., Vunjak-Novakovic G., Whitseh J. A., Morrisey E. E., The Cellular and Physiological Basis for Lung Repair and Regeneration: Past, Present, and Future. Cell stem cell 26, 482–502 (2020). - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous