

The palmitoylation of gasdermin D directs its membrane translocation and pore formation during pyroptosis
- PMID: 38530158
- PMCID: PMC11367861
- DOI: 10.1126/sciimmunol.adn1452
The palmitoylation of gasdermin D directs its membrane translocation and pore formation during pyroptosis
Abstract
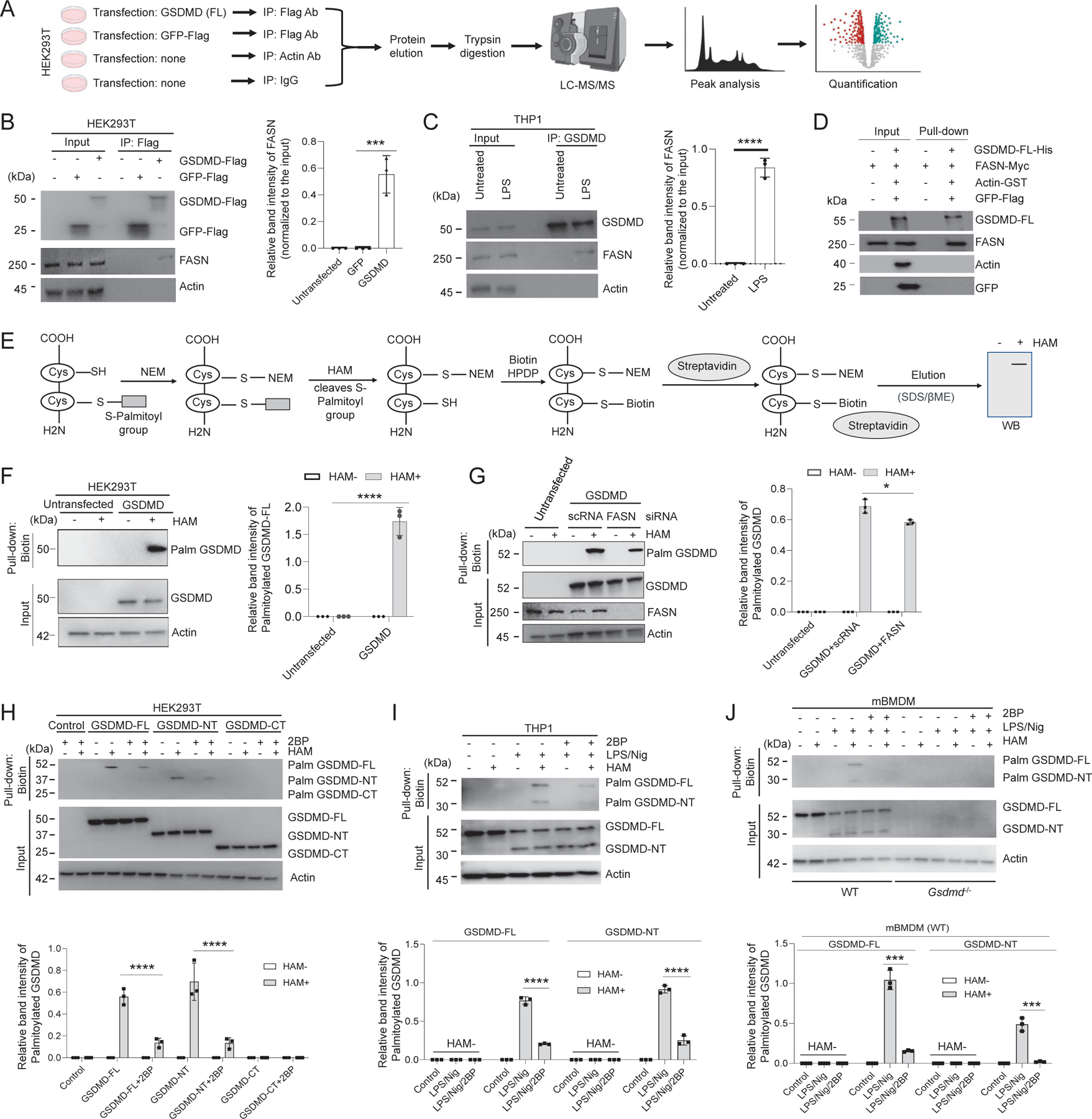
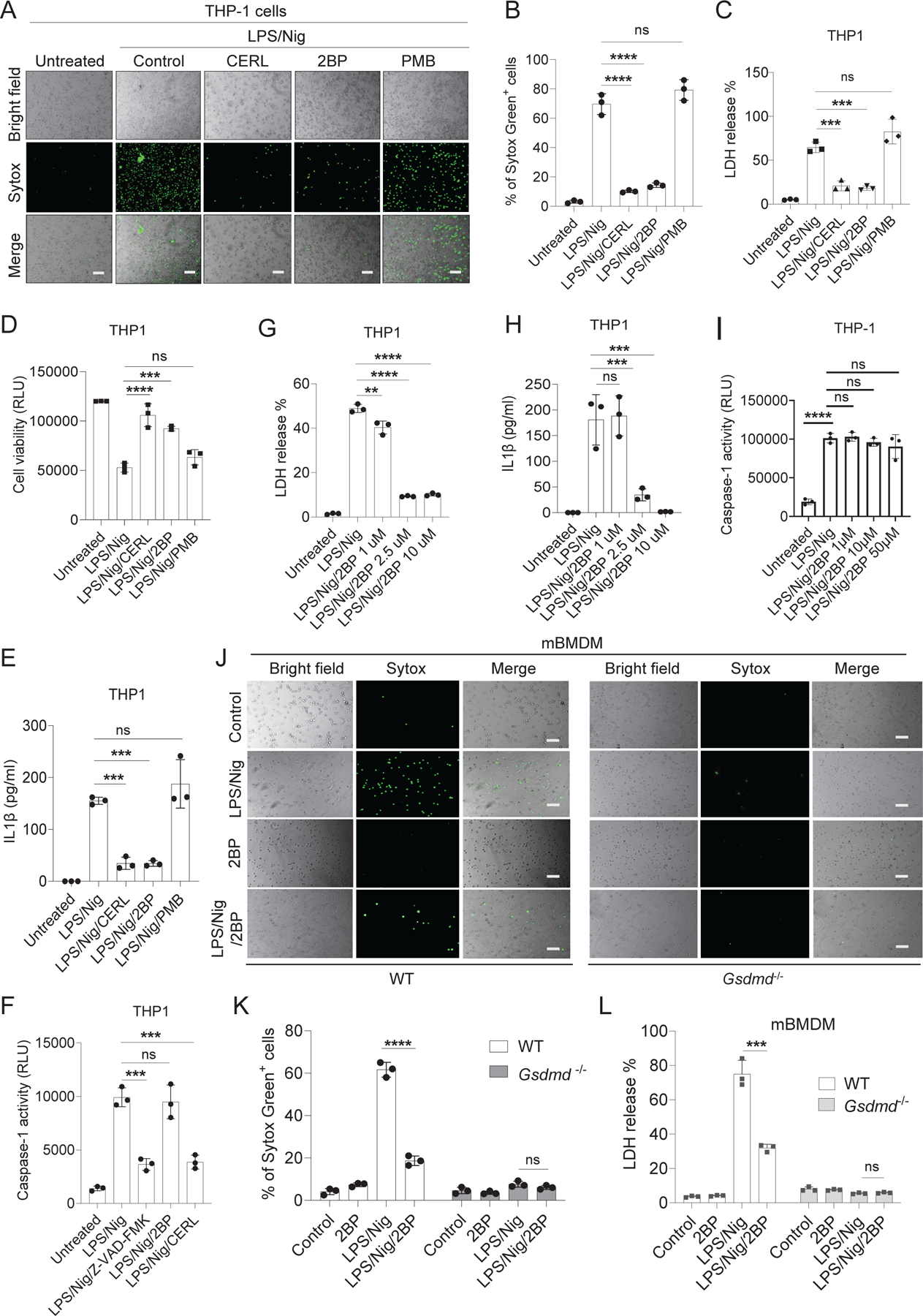
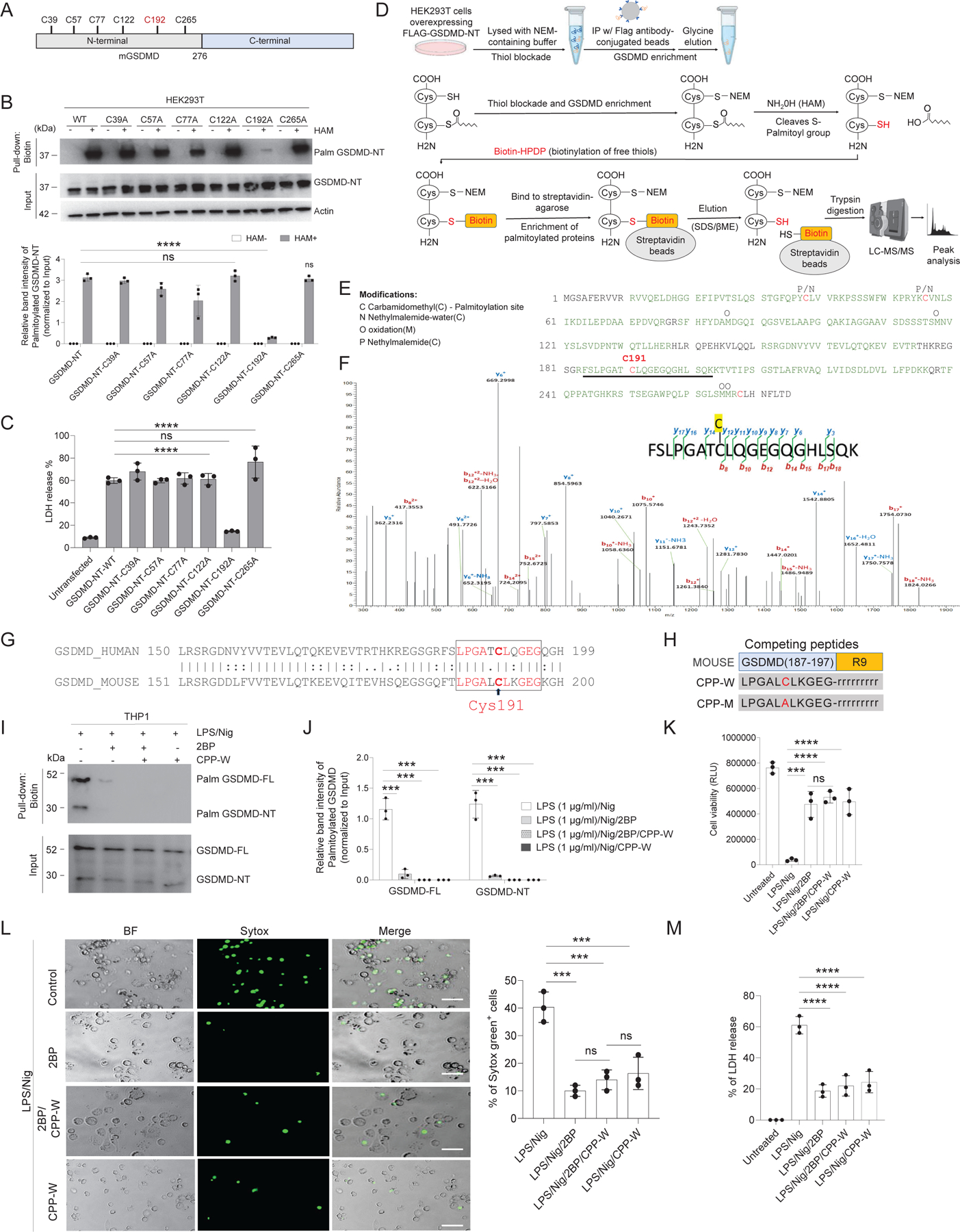
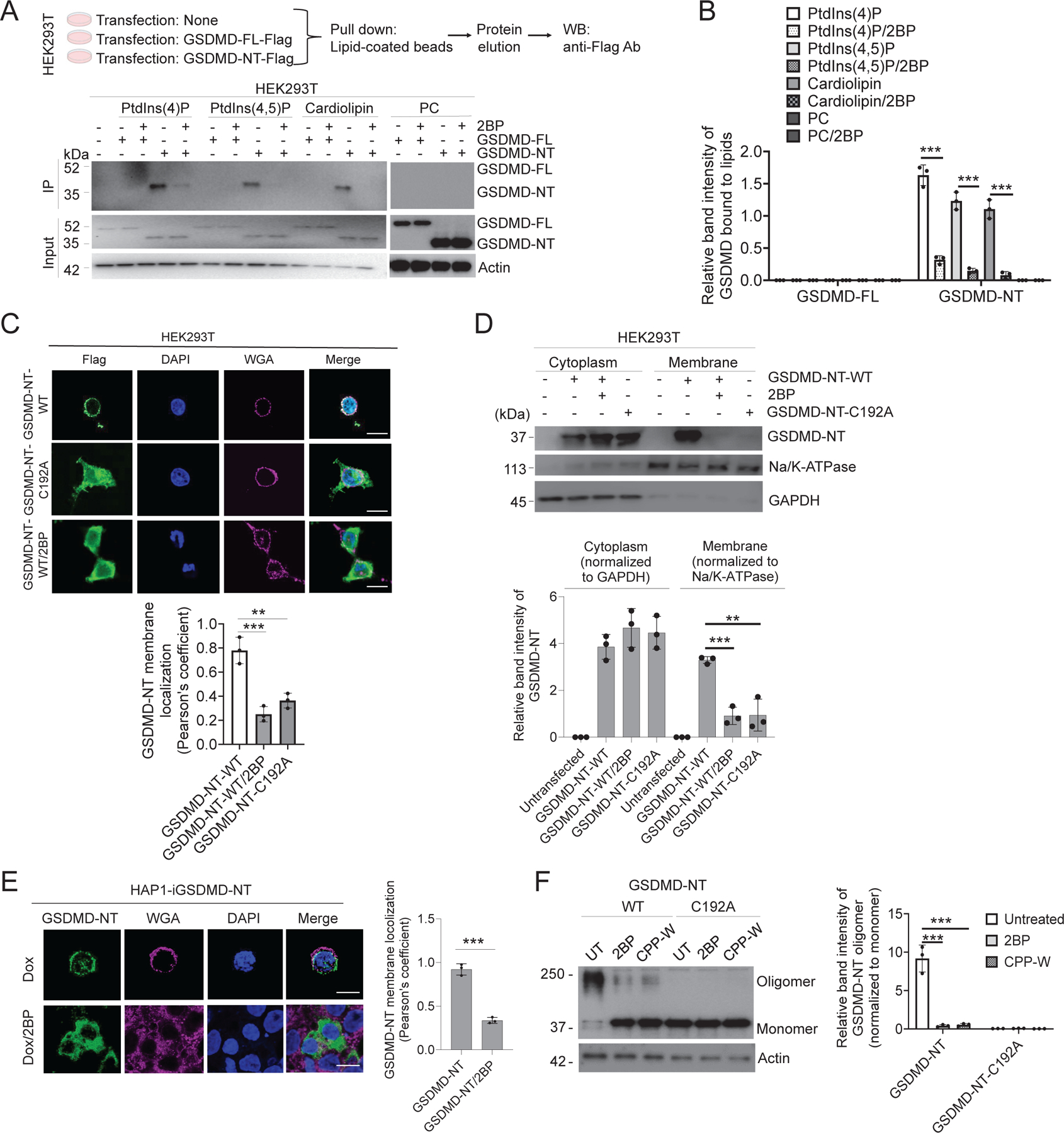
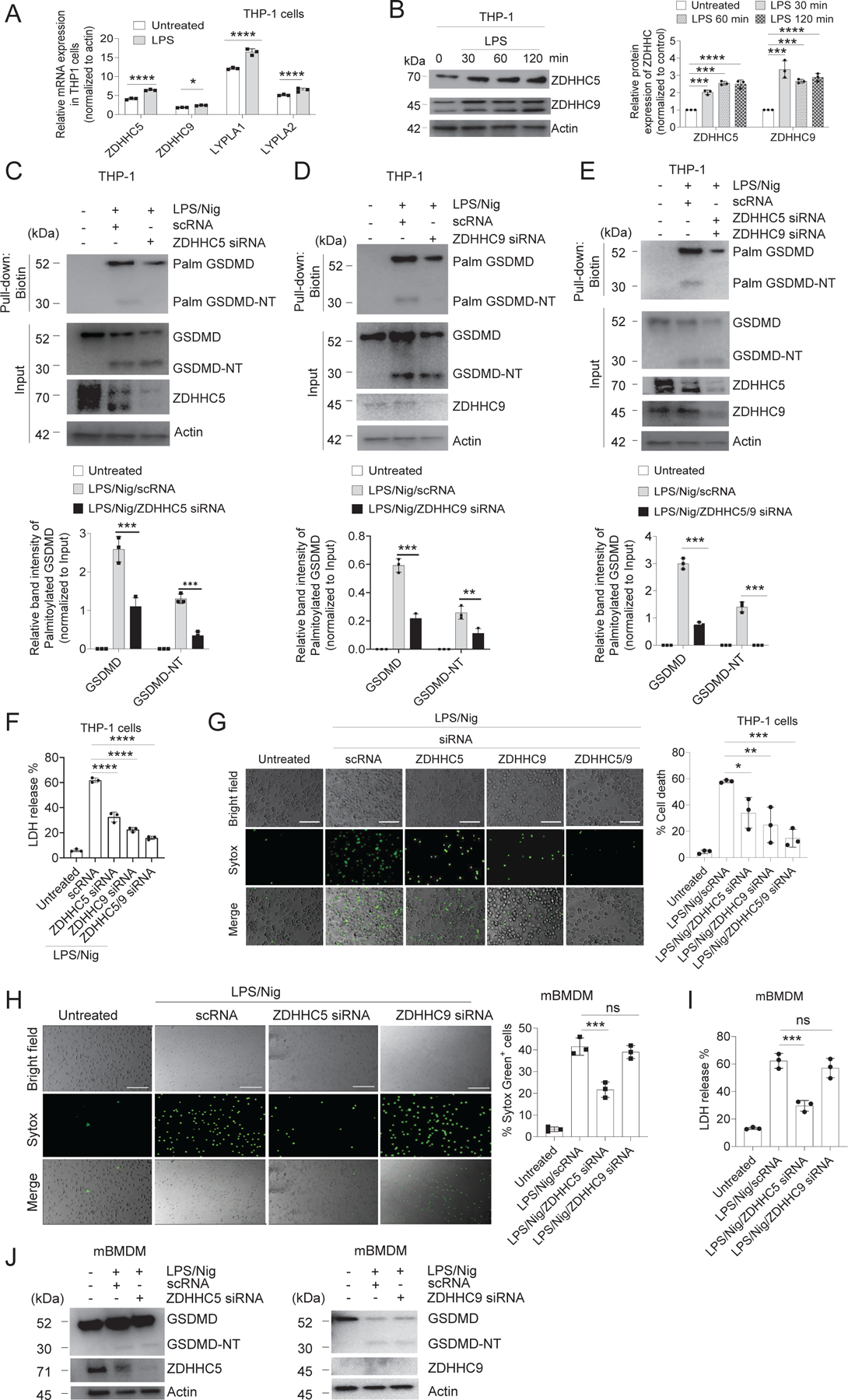
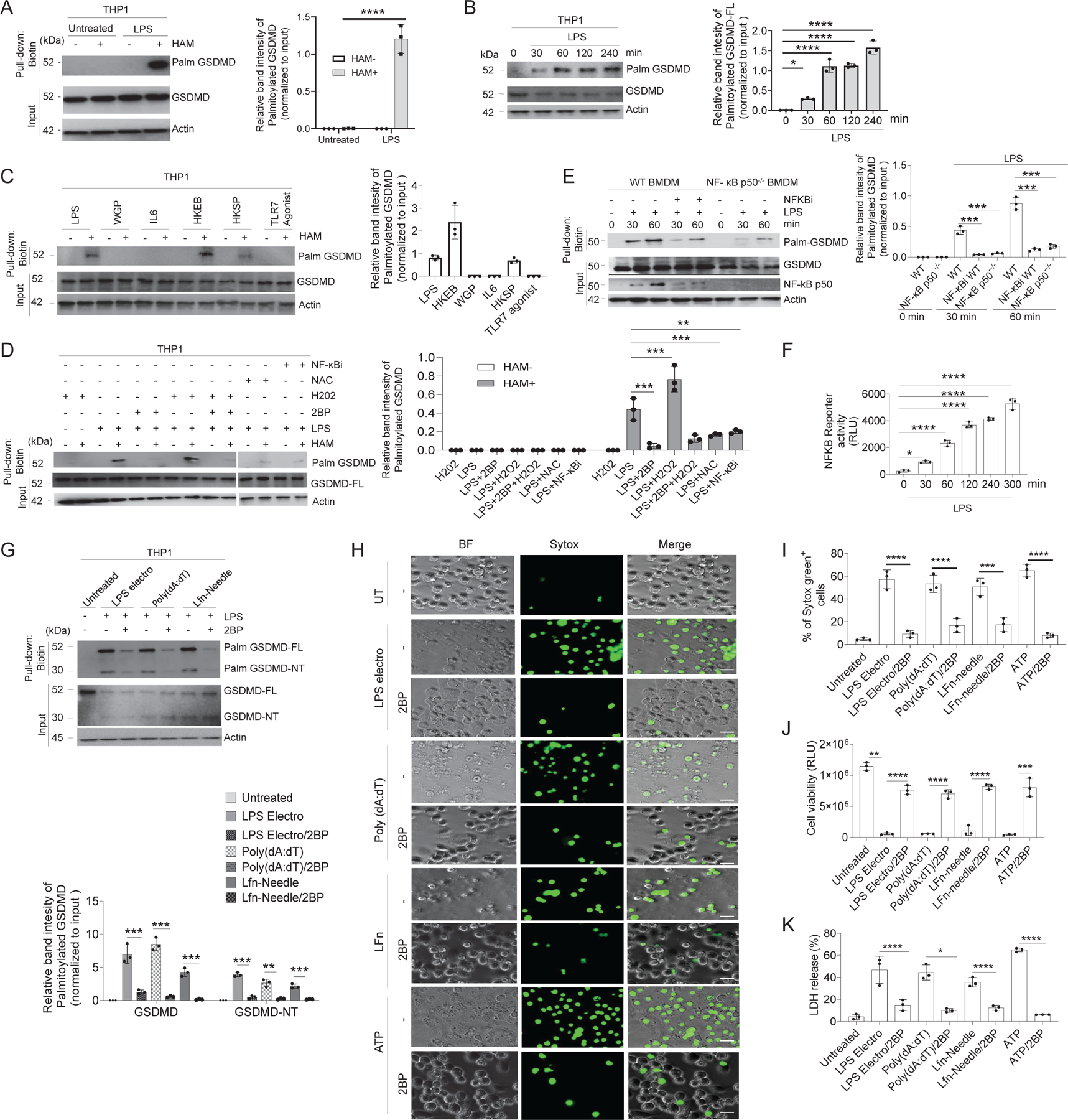
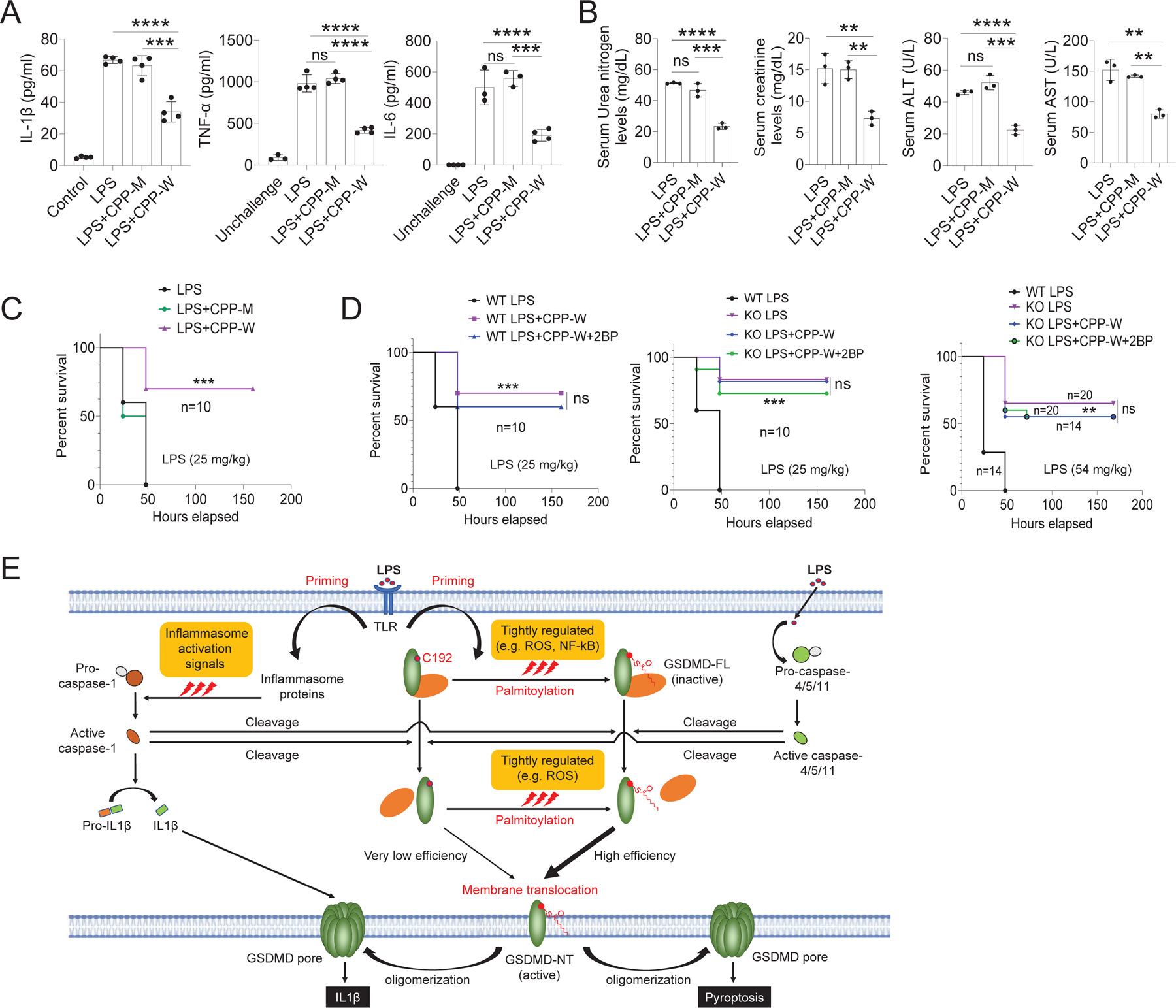
Plasma membrane perforation elicited by caspase cleavage of the gasdermin D (GSDMD) N-terminal domain (GSDMD-NT) triggers pyroptosis. The mechanisms underlying GSDMD membrane translocation and pore formation are not fully understood. Here, using a proteomic approach, we identified fatty acid synthase (FASN) as a GSDMD-binding partner. S-palmitoylation of GSDMD at Cys191/Cys192 (human/mouse), catalyzed by palmitoyl acyltransferases ZDHHC5 and ZDHHC9 and facilitated by reactive oxygen species (ROS), directly mediated membrane translocation of GSDMD-NT but not full-length GSDMD (GSDMD-FL). Palmitoylation of GSDMD-FL could be induced before inflammasome activation by stimuli such as lipopolysaccharide (LPS), consequently serving as an essential molecular event in macrophage priming. Inhibition of GSDMD palmitoylation suppressed macrophage pyroptosis and IL-1β release, mitigated organ damage, and enhanced the survival of septic mice. Thus, GSDMD-NT palmitoylation is a key regulatory mechanism controlling GSDMD membrane localization and activation, which may offer an additional target for modulating immune activity in infectious and inflammatory diseases.
Conflict of interest statement
Figures
Update of
-
Palmitoylation of gasdermin D directs its membrane translocation and pore formation in pyroptosis.bioRxiv [Preprint]. 2023 Feb 21:2023.02.21.529402. doi: 10.1101/2023.02.21.529402. bioRxiv. 2023. Update in: Sci Immunol. 2024 Apr 12;9(94):eadn1452. doi: 10.1126/sciimmunol.adn1452. PMID: 36865189 Free PMC article. Updated. Preprint.
References
-
- Shi J, Zhao Y, Wang K, Shi X, Wang Y, Huang H, Zhuang Y, Cai T, Wang F, Shao F, Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 526, 660–665 (2015). - PubMed
-
- Kayagaki N, Stowe IB, Lee BL, O’Rourke K, Anderson K, Warming S, Cuellar T, Haley B, Roose-Girma M, Phung QT, Liu PS, Lill JR, Li H, Wu J, Kummerfeld S, Zhang J, Lee WP, Snipas SJ, Salvesen GS, Morris LX, Fitzgerald L, Zhang Y, Bertram EM, Goodnow CC, Dixit VM, Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 526, 666–671 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01 HL158688/HL/NHLBI NIH HHS/United States
- T32 HL066987/HL/NHLBI NIH HHS/United States
- R01 AI141386/AI/NIAID NIH HHS/United States
- S10 OD030322/OD/NIH HHS/United States
- DP1 HD087988/HD/NICHD NIH HHS/United States
- P30 DK034854/DK/NIDDK NIH HHS/United States
- U19 AI133524/AI/NIAID NIH HHS/United States
- R01 HL092020/HL/NHLBI NIH HHS/United States
- R01 AI139914/AI/NIAID NIH HHS/United States
- R01 AI116550/AI/NIAID NIH HHS/United States
- R01 AI145274/AI/NIAID NIH HHS/United States
- P50 HD105351/HD/NICHD NIH HHS/United States
- R01 AI124491/AI/NIAID NIH HHS/United States
- R01 AI142642/AI/NIAID NIH HHS/United States
- R37 AI116550/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
