

Sequence Dependence in Nucleosome Dynamics
- PMID: 38530903
- PMCID: PMC11181342
- DOI: 10.1021/acs.jpcb.3c07363
Sequence Dependence in Nucleosome Dynamics
Abstract
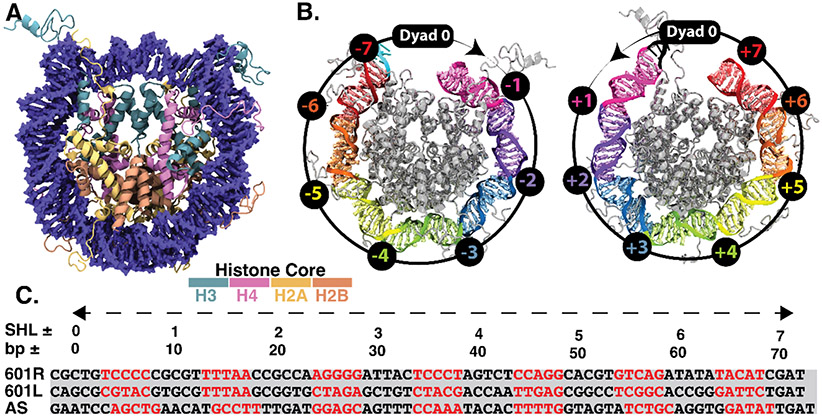
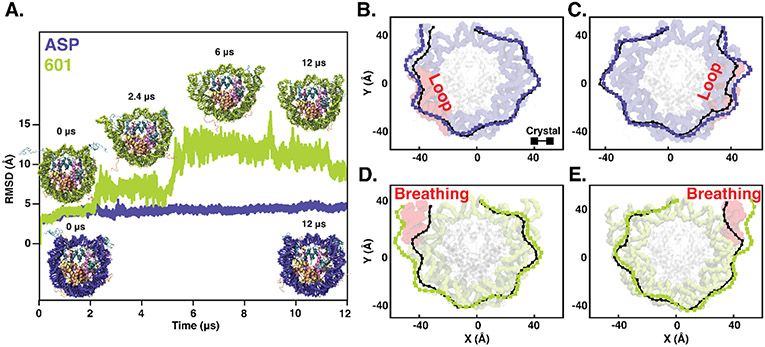
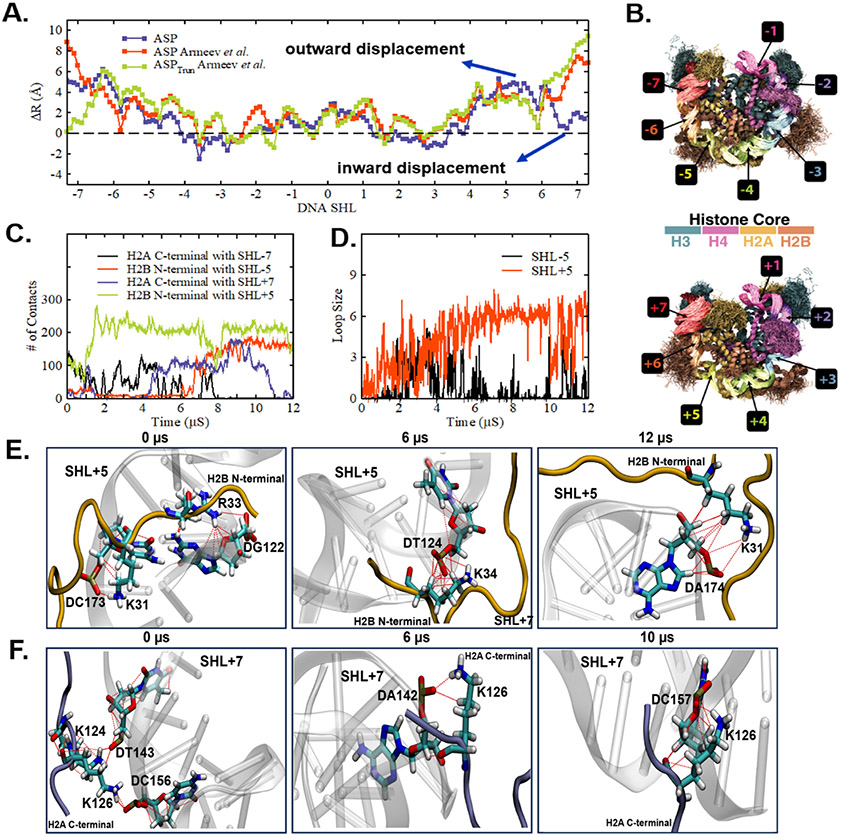
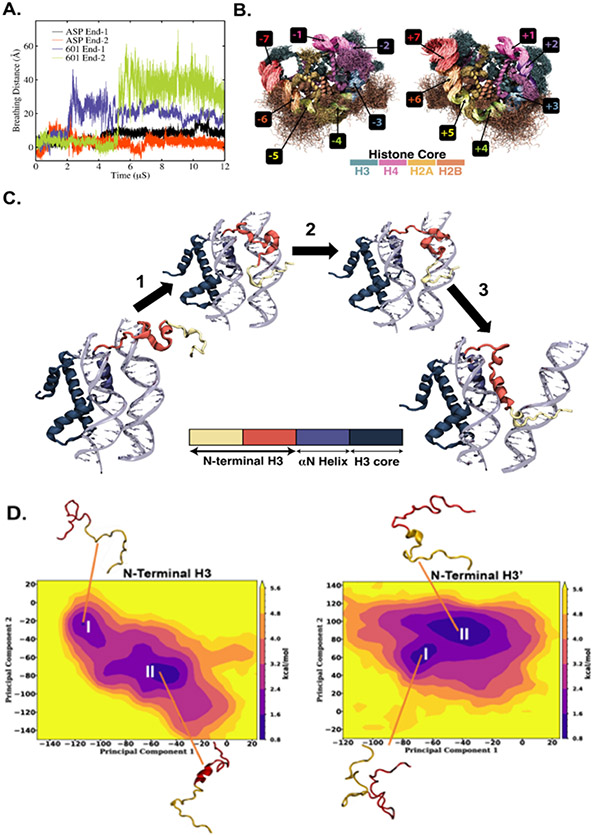
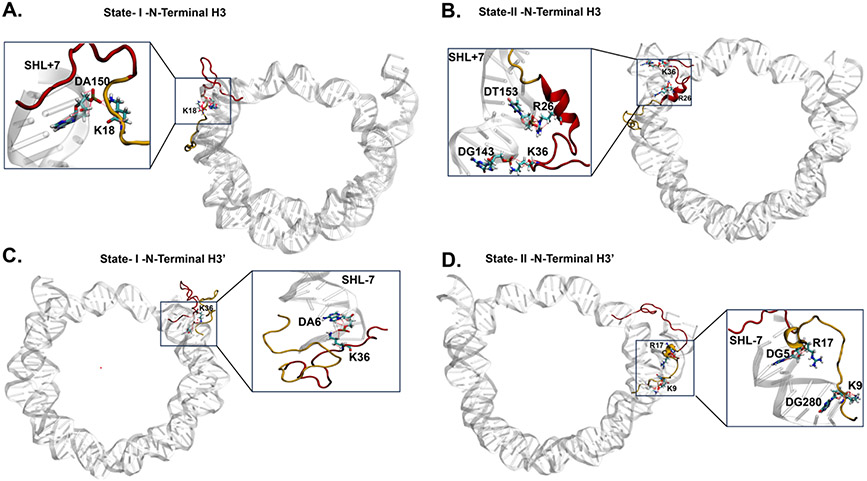
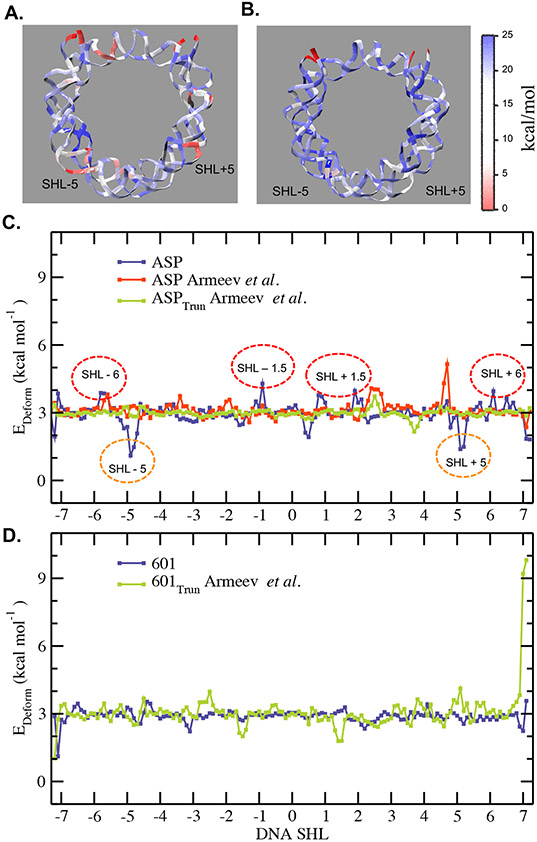
The basic packaging unit of eukaryotic chromatin is the nucleosome that contains 145-147 base pair duplex DNA wrapped around an octameric histone protein. While the DNA sequence plays a crucial role in controlling the positioning of the nucleosome, the molecular details behind the interplay between DNA sequence and nucleosome dynamics remain relatively unexplored. This study analyzes this interplay in detail by performing all-atom molecular dynamics simulations of nucleosomes, comparing the human α-satellite palindromic (ASP) and the strong positioning "Widom-601" DNA sequence at time scales of 12 μs. The simulations are performed at salt concentrations 10-20 times higher than physiological salt concentrations to screen the electrostatic interactions and promote unwrapping. These microsecond-long simulations give insight into the molecular-level sequence-dependent events that dictate the pathway of DNA unwrapping. We find that the "ASP" sequence forms a loop around SHL ± 5 for three sets of simulations. Coincident with loop formation is a cooperative increase in contacts with the neighboring N-terminal H2B tail and C-terminal H2A tail and the release of neighboring counterions. We find that the Widom-601 sequence exhibits a strong breathing motion of the nucleic acid ends. Coincident with the breathing motion is the collapse of the full N-terminal H3 tail and formation of an α-helix that interacts with the H3 histone core. We postulate that the dynamics of these histone tails and their modification with post-translational modifications (PTMs) may play a key role in governing this dynamics.
Figures
References
-
- Kornberg RD; Lorch Y Twenty-five years of the nucleosome, fundamental particle of the eukaryote chromosome. Cell 1999, 98 (3), 285–294. - PubMed
-
- Davey CA; Sargent DF; Luger K; Maeder AW; Richmond TJ Solvent mediated interactions in the structure of the nucleosome core particle at 1.9 å resolution. Journal of molecular biology 2002, 319 (5), 1097–1113. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
