

Ancestry dynamics and trait selection in a designer cat breed
- PMID: 38531359
- PMCID: PMC11162505
- DOI: 10.1016/j.cub.2024.02.075
Ancestry dynamics and trait selection in a designer cat breed
Abstract
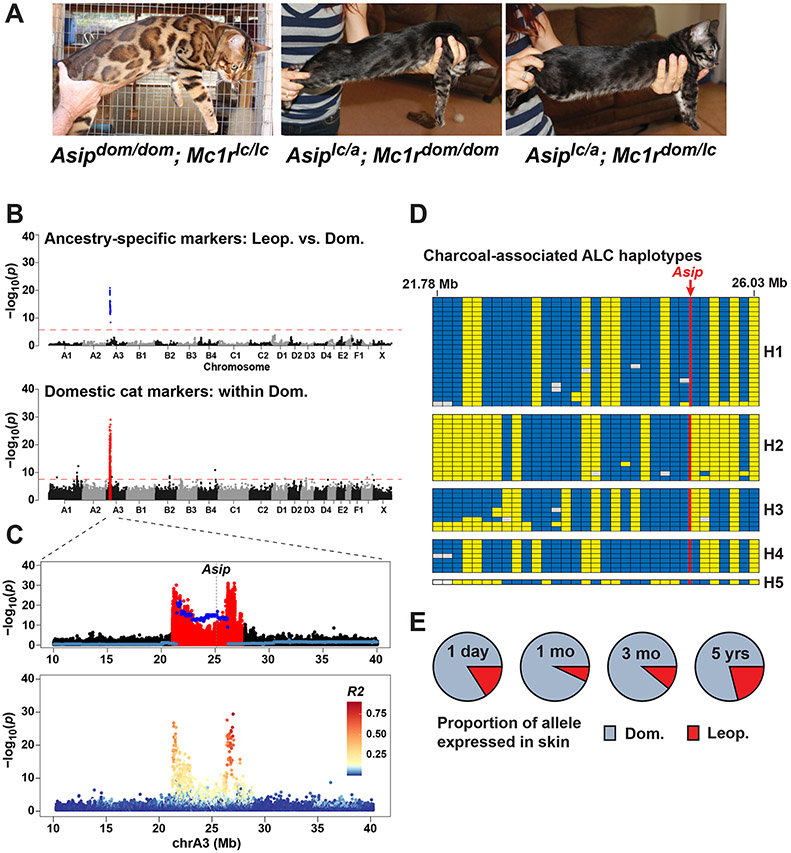
The Bengal cat breed was developed from intercrosses between the Asian leopard cat, Prionailurus bengalensis, and the domestic cat, Felis catus, with a last common ancestor approximately 6 million years ago. Predicted to derive ∼94% of their genome from domestic cats, regions of the leopard cat genome are thought to account for the unique pelage traits and ornate color patterns of the Bengal breed, which are similar to those of ocelots and jaguars. We explore ancestry distribution and selection signatures in the Bengal breed by using reduced representation and whole-genome sequencing from 947 cats. The mean proportion of leopard cat DNA in the Bengal breed is 3.48%, lower than predicted from breed history, and is broadly distributed, covering 93% of the Bengal genome. Overall, leopard cat introgressions do not show strong signatures of selection across the Bengal breed. However, two popular color traits in Bengal cats, charcoal and pheomelanin intensity, are explained by selection of leopard cat genes whose expression is reduced in a domestic cat background, consistent with genetic incompatibility resulting from hybridization. We characterize several selective sweeps in the Bengal genome that harbor candidate genes for pelage and color pattern and that are associated with domestic, rather than leopard, cat haplotypes. We identify the molecular and phenotypic basis of one selective sweep as reduced expression of the Fgfr2 gene, which underlies glitter, a trait desired by breeders that affects hair texture and light reflectivity.
Keywords: genetic incompatibility; genome-wide association; interspecies hybridization; introgression; pigmentation pathways.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






References
-
- TICA (2019). The Bengal Breed, https://www.tica.org/breeds/browse-all-breeds?view=article&id=1521:the-b....
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
