

METTL3-mediated m6A modification increases Hspa1a stability to inhibit osteoblast aging
- PMID: 38538596
- PMCID: PMC10973419
- DOI: 10.1038/s41420-024-01925-4
METTL3-mediated m6A modification increases Hspa1a stability to inhibit osteoblast aging
Abstract
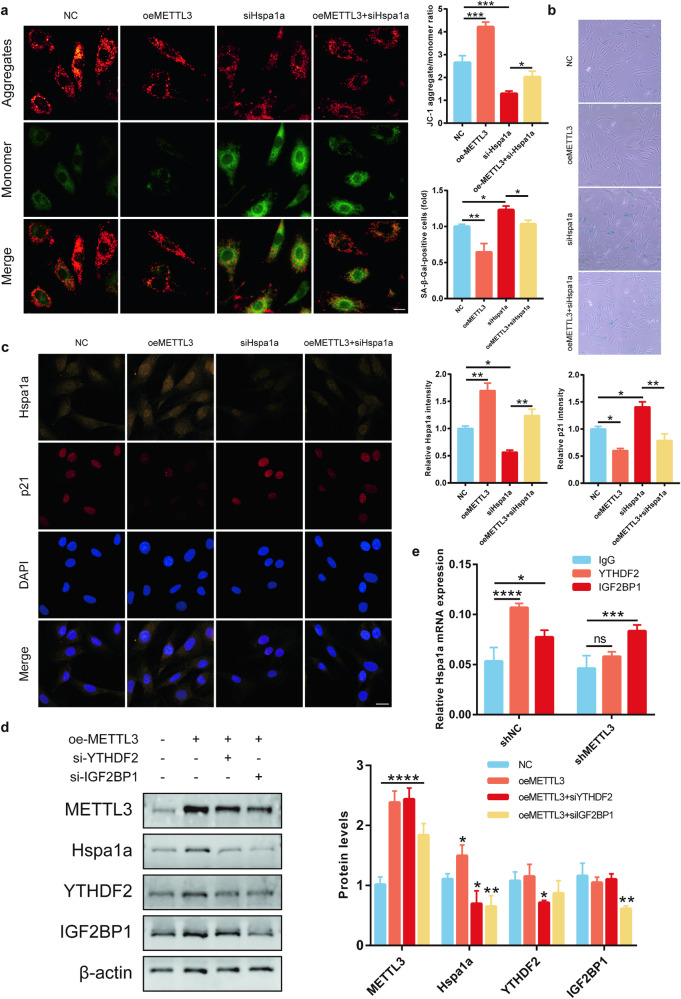
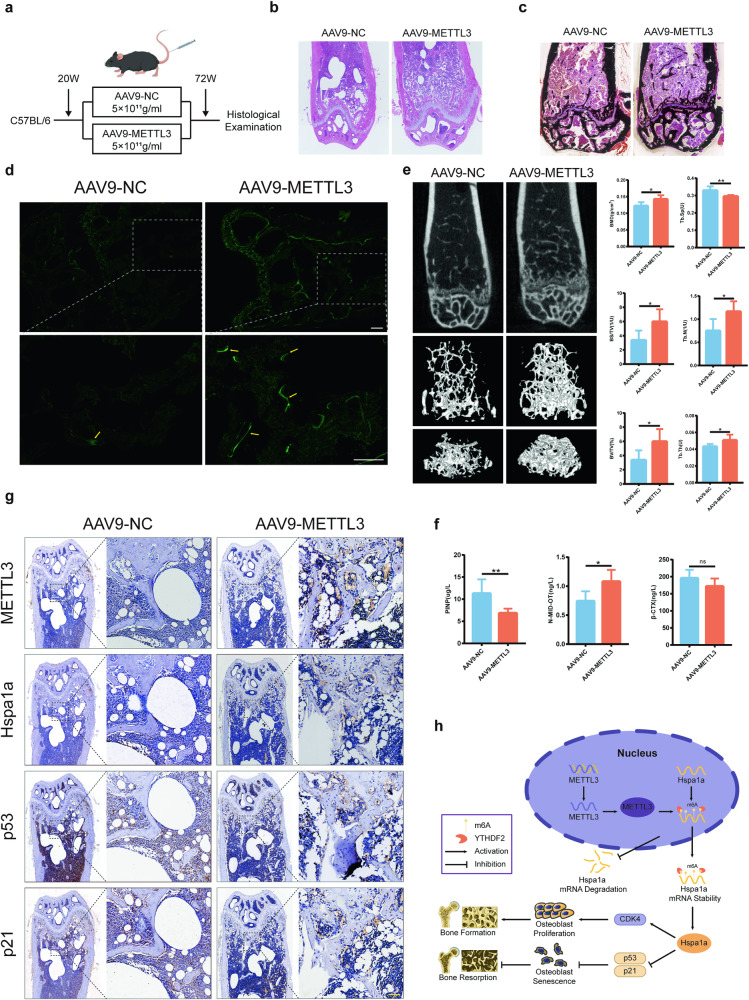
Senile osteoporosis is mainly caused by osteoblasts attenuation, which results in reduced bone mass and disrupted bone remodeling. Numerous studies have focused on the regulatory role of m6A modification in osteoporosis; however, most of the studies have investigated the differentiation of bone marrow mesenchymal stem cells (BMSCs), while the direct regulatory mechanism of m6A on osteoblasts remains unknown. This study revealed that the progression of senile osteoporosis is closely related to the downregulation of m6A modification and methyltransferase-like 3 (METTL3). Overexpression of METTL3 inhibits osteoblast aging. Methylated RNA immunoprecipitation sequencing (MeRIP-seq) revealed that METTL3 upregulates the stability of Hspa1a mRNA, thereby inhibiting osteoblast aging. Moreover, the results demonstrated that METTL3 enhances the stability of Hspa1a mRNA via m6A modification to regulate osteoblast aging. Notably, YTH N6-methyladenosine RNA binding protein 2 (YTHDF2) participates in stabilizing Hspa1a mRNA in the METTL3-mediated m6A modification process, rather than the well-known degradation function. Mechanistically, METTL3 increases the stability of Hspa1a mRNA in a YTHDF2-dependent manner to inhibit osteoblast aging. Our results confirmed the significant role of METTL3 in osteoblast aging and suggested that METTL3 could be a potential therapeutic target for senile osteoporosis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Grants and funding
- 82060405/National Natural Science Foundation of China (National Science Foundation of China)
- 82360436/National Natural Science Foundation of China (National Science Foundation of China)
- 81960403/National Natural Science Foundation of China (National Science Foundation of China)
- 2023CXZX-157/Gansu Education Department (Education Department of Gansu Province)
LinkOut - more resources
Full Text Sources
