

De novo GRIN variants in M3 helix associated with neurological disorders control channel gating of NMDA receptor
- PMID: 38538865
- PMCID: PMC10973091
- DOI: 10.1007/s00018-023-05069-z
De novo GRIN variants in M3 helix associated with neurological disorders control channel gating of NMDA receptor
Abstract
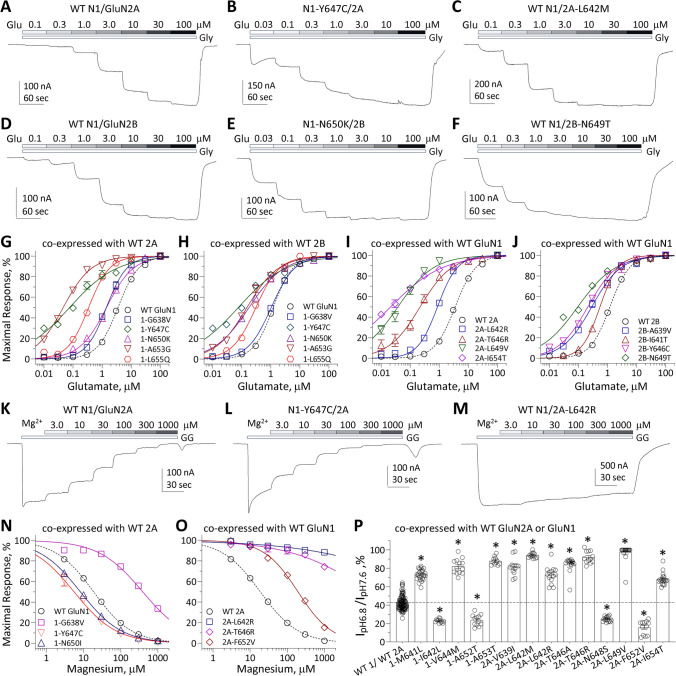
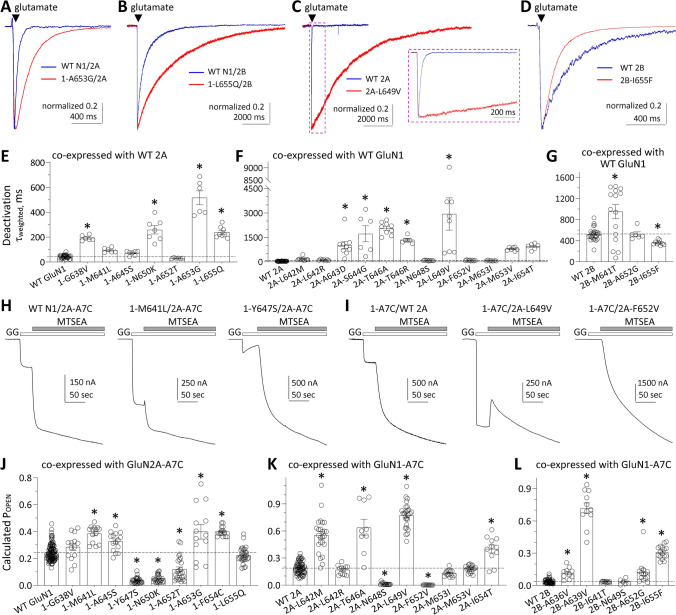
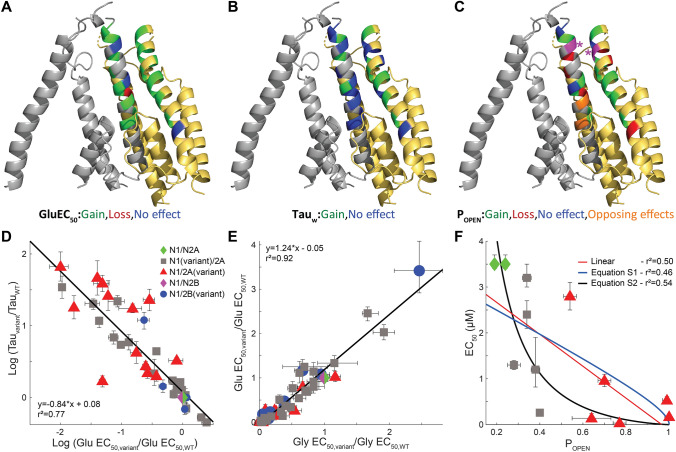
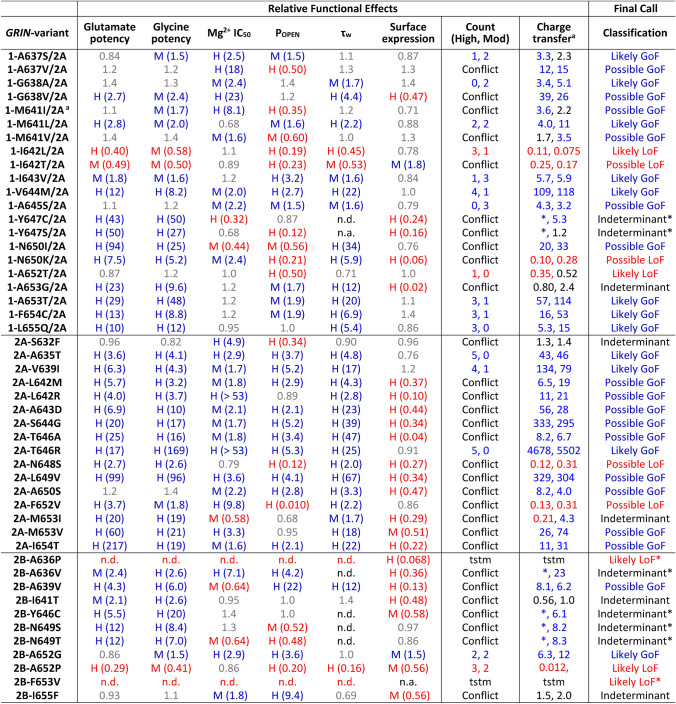
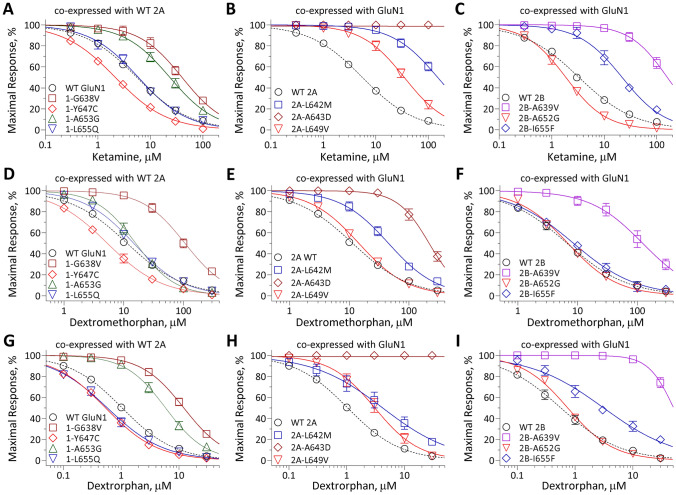
N-methyl-D-aspartate receptors (NMDARs) are members of the glutamate receptor family and participate in excitatory postsynaptic transmission throughout the central nervous system. Genetic variants in GRIN genes encoding NMDAR subunits are associated with a spectrum of neurological disorders. The M3 transmembrane helices of the NMDAR couple directly to the agonist-binding domains and form a helical bundle crossing in the closed receptors that occludes the pore. The M3 functions as a transduction element whose conformational change couples ligand binding to opening of an ion conducting pore. In this study, we report the functional consequences of 48 de novo missense variants in GRIN1, GRIN2A, and GRIN2B that alter residues in the M3 transmembrane helix. These de novo variants were identified in children with neurological and neuropsychiatric disorders including epilepsy, developmental delay, intellectual disability, hypotonia and attention deficit hyperactivity disorder. All 48 variants in M3 for which comprehensive testing was completed produce a gain-of-function (28/48) compared to loss-of-function (9/48); 11 variants had an indeterminant phenotype. This supports the idea that a key structural feature of the M3 gate exists to stabilize the closed state so that agonist binding can drive channel opening. Given that most M3 variants enhance channel gating, we assessed the potency of FDA-approved NMDAR channel blockers on these variant receptors. These data provide new insight into the structure-function relationship of the NMDAR gate, and suggest that variants within the M3 transmembrane helix produce a gain-of-function.
Keywords: Channelopathy; Functional genomics; Glutamate receptor; NMDA receptor; TMD.
© 2024. The Author(s).
Conflict of interest statement
SFT is a member of the SAB for Sage Therapeutics, Eumentis Therapeutics, the GRIN2B Foundation, the CureGRIN Foundation, and CombinedBrain. SFT is consultant for GRIN Therapeutics and Neurocrine, a cofounder of NeurOp, Inc. and Agrithera, and a member of the Board of Directors for NeurOp Inc. HY is the PI on a research grant from Sage Therapeutics to Emory and SJM is PI on a grant from GRIN Therapeutics to Emory. TAB—Consultancy for AveXis, Ovid, GW Pharmaceuticals, International Rett Syndrome Foundation, Takeda, Taysha, CureGRIN, GRIN Therapeutics, Alcyone, Neurogene, and Marinus; Clinical Trials with Acadia, Ovid, GW Pharmaceuticals, Marinus and RSRT; all remuneration has been made to his department.
Figures


References
-
- Hansen KB, Wollmuth LP, Bowie D, Furukawa H, Menniti FS, Sobolevsky AI, Swanson GT, Swanger SA, Greger IH, Nakagawa T, McBain CJ, Jayaraman V, Low CM, Dell'Acqua ML, Diamond JS, Camp CR, Perszyk RE, Yuan H, Traynelis SF. Structure, function, and pharmacology of glutamate receptor ion channels. Pharmacol Rev. 2021;73(4):298–487. doi: 10.1124/pharmrev.120.000131. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- R01 AG075444/AG/NIA NIH HHS/United States
- K02 NS112600/NS/NINDS NIH HHS/United States
- R01 NS131512/NS/NINDS NIH HHS/United States
- AG081401/AG/NIA NIH HHS/United States
- R01 HD082373/HD/NICHD NIH HHS/United States
- R21 AG072142/AG/NIA NIH HHS/United States
- AG079956/AG/NIA NIH HHS/United States
- R01 AG079956/AG/NIA NIH HHS/United States
- NS111619/NS/NINDS NIH HHS/United States
- AG072142/AG/NIA NIH HHS/United States
- AG075444/AG/NIA NIH HHS/United States
- R35 NS111619/NS/NINDS NIH HHS/United States
- R21 MH127404/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
