

Data integration strategies for whole-cell modeling
- PMID: 38544322
- PMCID: PMC11042497
- DOI: 10.1093/femsyr/foae011
Data integration strategies for whole-cell modeling
Abstract
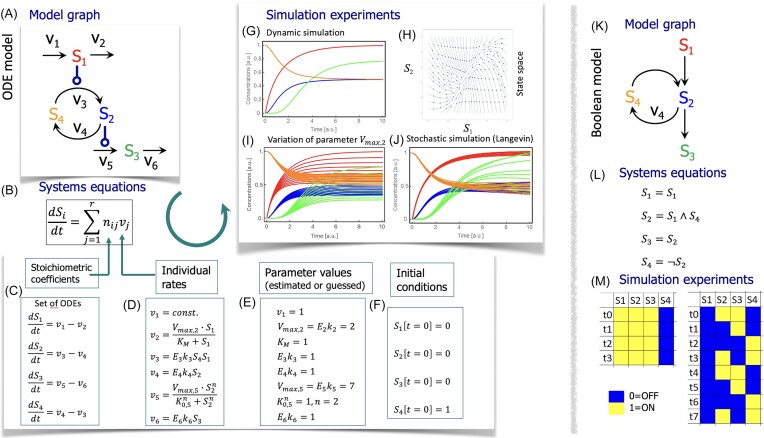
Data makes the world go round-and high quality data is a prerequisite for precise models, especially for whole-cell models (WCM). Data for WCM must be reusable, contain information about the exact experimental background, and should-in its entirety-cover all relevant processes in the cell. Here, we review basic requirements to data for WCM and strategies how to combine them. As a species-specific resource, we introduce the Yeast Cell Model Data Base (YCMDB) to illustrate requirements and solutions. We discuss recent standards for data as well as for computational models including the modeling process as data to be reported. We outline strategies for constructions of WCM despite their inherent complexity.
Keywords: complex networks; emerging standards; single cell; yeast cell model.
© The Author(s) 2024. Published by Oxford University Press on behalf of FEMS.
Conflict of interest statement
None declared.
Figures




References
-
- Adrover MA, Zi Z, Duch A et al. Time-dependent quantitative multicomponent control of the G1-S network by the stress-activated protein kinase Hog1 upon osmostress. Sci Signal. 2011;4:ra63. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
