

Allele-specific DNA methylation and gene expression during shoot organogenesis in tissue culture of hybrid poplar
- PMID: 38544548
- PMCID: PMC10967691
- DOI: 10.1093/hr/uhae027
Allele-specific DNA methylation and gene expression during shoot organogenesis in tissue culture of hybrid poplar
Abstract
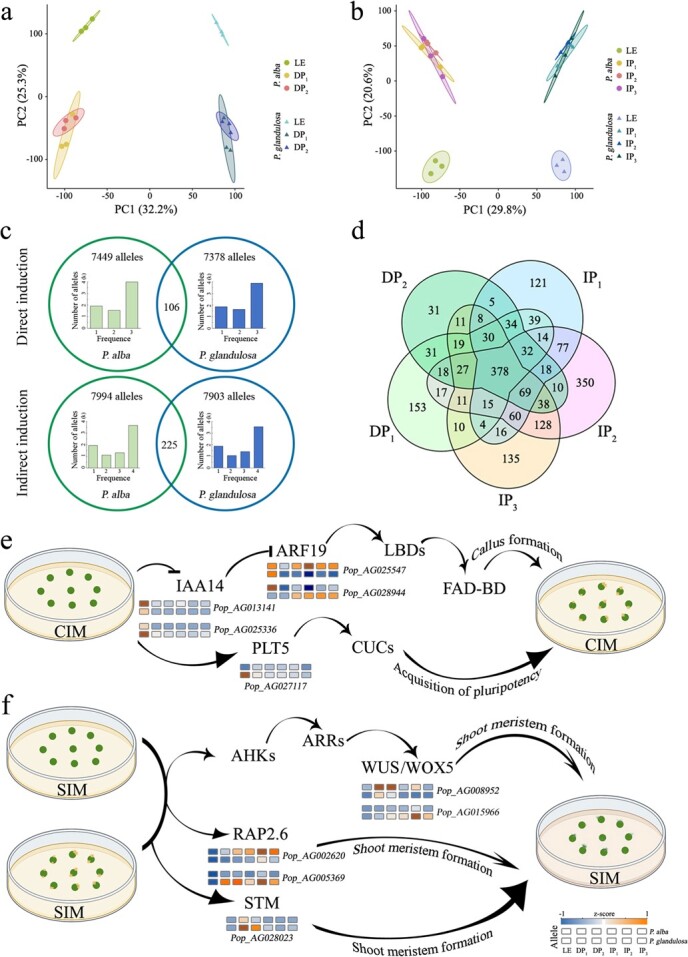
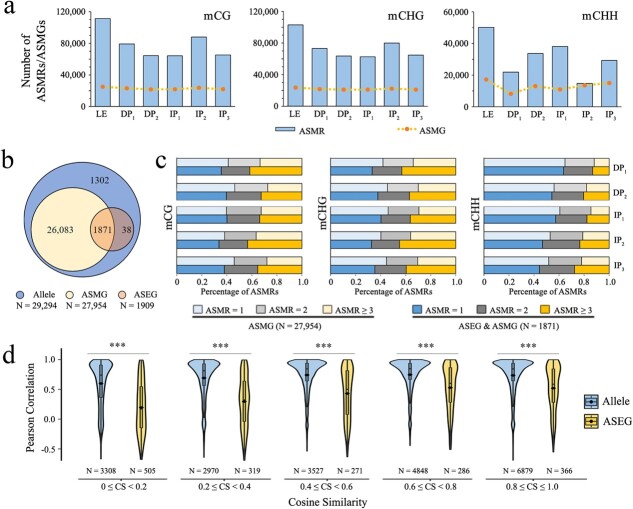
Plant tissue regeneration is critical for genetic transformation and genome editing techniques. During the regeneration process, changes in epigenetic modifications accompany the cell fate transition. However, how allele-specific DNA methylation in two haplotypes contributes to the transcriptional dynamics during regeneration remains elusive. Here we applied an inter-species hybrid poplar (Populus alba × P. glandulosa cv. 84 K) as a system to characterize the DNA methylation landscape during de novo shoot organogenesis at allele level. Both direct and indirect shoot organogenesis showed a reduction in genome-wide DNA methylation. At gene level, non-expressed genes were hypermethylated in comparison with expressed genes. Among the genes exhibiting significant correlations between levels of DNA methylation and gene expression, the expression patterns of 75% of genes were negatively correlated with DNA methylation in the CG context, whereas the correlation patterns in the CHH context were the reverse. The allele-biased DNA methylation was consistent during shoot organogenesis, with fewer than one-thousandth of allele-specific methylation regions shifted. Analysis of allele-specific expression revealed that there were only 1909 genes showing phase-dependent allele-biased expression in the regeneration process, among which the allele pairs with greater differences in transcription factor binding sites at promoter regions exhibited greater differences in allele expression. Our results indicated a relatively independent transcriptional regulation in two subgenomes during shoot organogenesis, which was contributed by cis-acting genomic and epigenomic variations.
© The Author(s) 2024. Published by Oxford University Press on behalf of Nanjing Agricultural University.
Figures



References
-
- Ikeuchi M, Favero DS, Sakamoto Y. et al. . Molecular mechanisms of plant regeneration. Annu Rev Plant Biol. 2019;70:377–406 - PubMed
-
- Atta R, Laurens L, Boucheron-Dubuisson E. et al. . Pluripotency of Arabidopsis xylem pericycle underlies shoot regeneration from root and hypocotyl explants grown in vitro. Plant J. 2009;57:626–44 - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
