

Crimean-Congo haemorrhagic fever virus uses LDLR to bind and enter host cells
- PMID: 38548922
- PMCID: PMC11153131
- DOI: 10.1038/s41564-024-01672-3
Crimean-Congo haemorrhagic fever virus uses LDLR to bind and enter host cells
Abstract
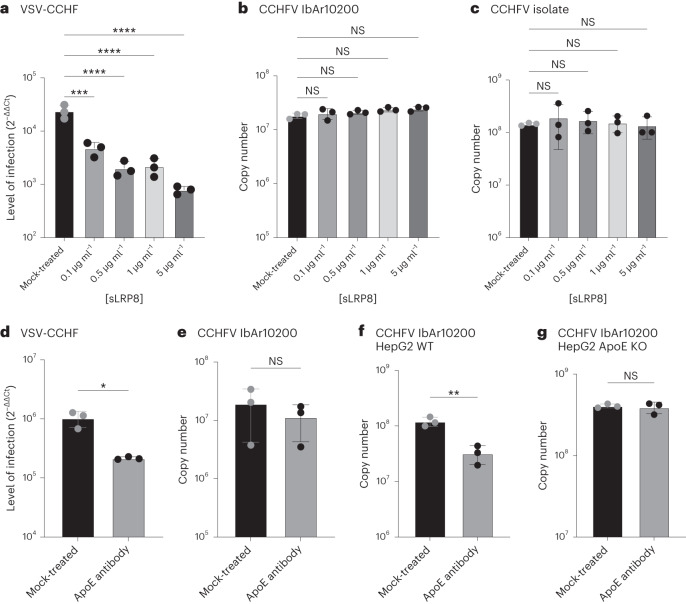
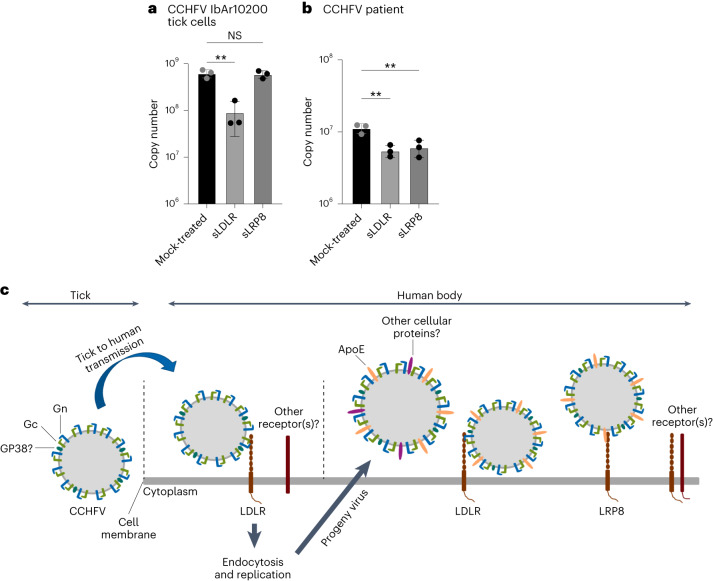
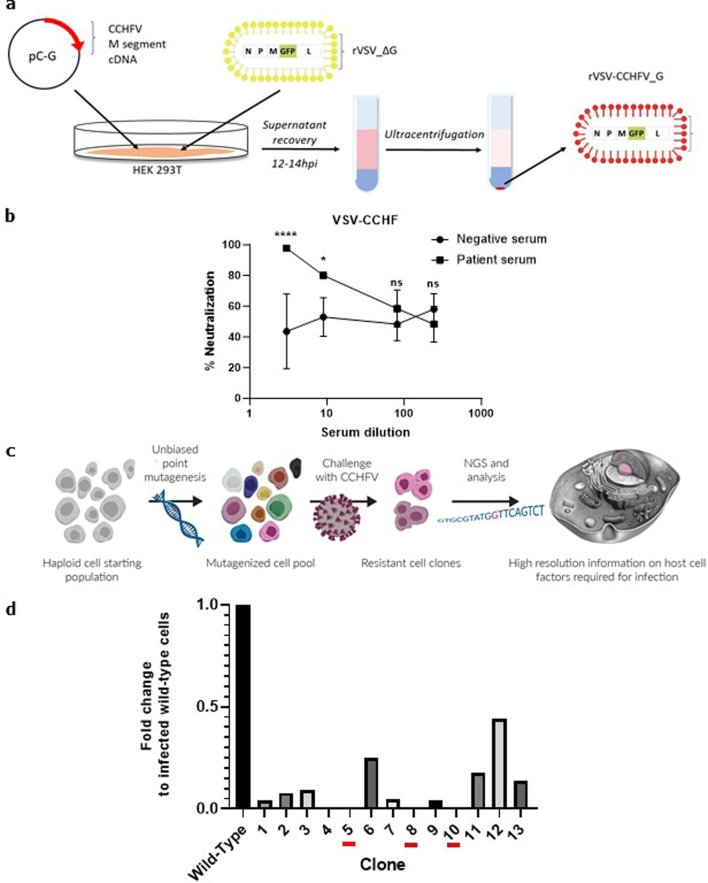
Climate change and population densities accelerated transmission of highly pathogenic viruses to humans, including the Crimean-Congo haemorrhagic fever virus (CCHFV). Here we report that the Low Density Lipoprotein Receptor (LDLR) is a critical receptor for CCHFV cell entry, playing a vital role in CCHFV infection in cell culture and blood vessel organoids. The interaction between CCHFV and LDLR is highly specific, with other members of the LDLR protein family failing to bind to or neutralize the virus. Biosensor experiments demonstrate that LDLR specifically binds the surface glycoproteins of CCHFV. Importantly, mice lacking LDLR exhibit a delay in CCHFV-induced disease. Furthermore, we identified the presence of Apolipoprotein E (ApoE) on CCHFV particles. Our findings highlight the essential role of LDLR in CCHFV infection, irrespective of ApoE presence, when the virus is produced in tick cells. This discovery holds profound implications for the development of future therapies against CCHFV.
© 2024. The Author(s).
Conflict of interest statement
J.M.P. is a founder and shareholder of JLP. V.M.L. is co‐founder, CEO and shareholder of HepaPredict AB, as well as co-founder and shareholder of PersoMedix AB, and discloses consultancy work for Enginzyme AB. M.H. is co‐founder, CEO and shareholder of Acus Laboratories GmbH and CSO of JLP Health GmbH. M.D. is an employee of Acus Laboratories GmbH and JLP Health GmbH. A.I. is an employee of Attana AB and T.A. is CSO of Attana AB. A patent application has been filed (PCT application, 2023, European patent EP 23 174 811.2). All other authors declare no competing interests.
Figures
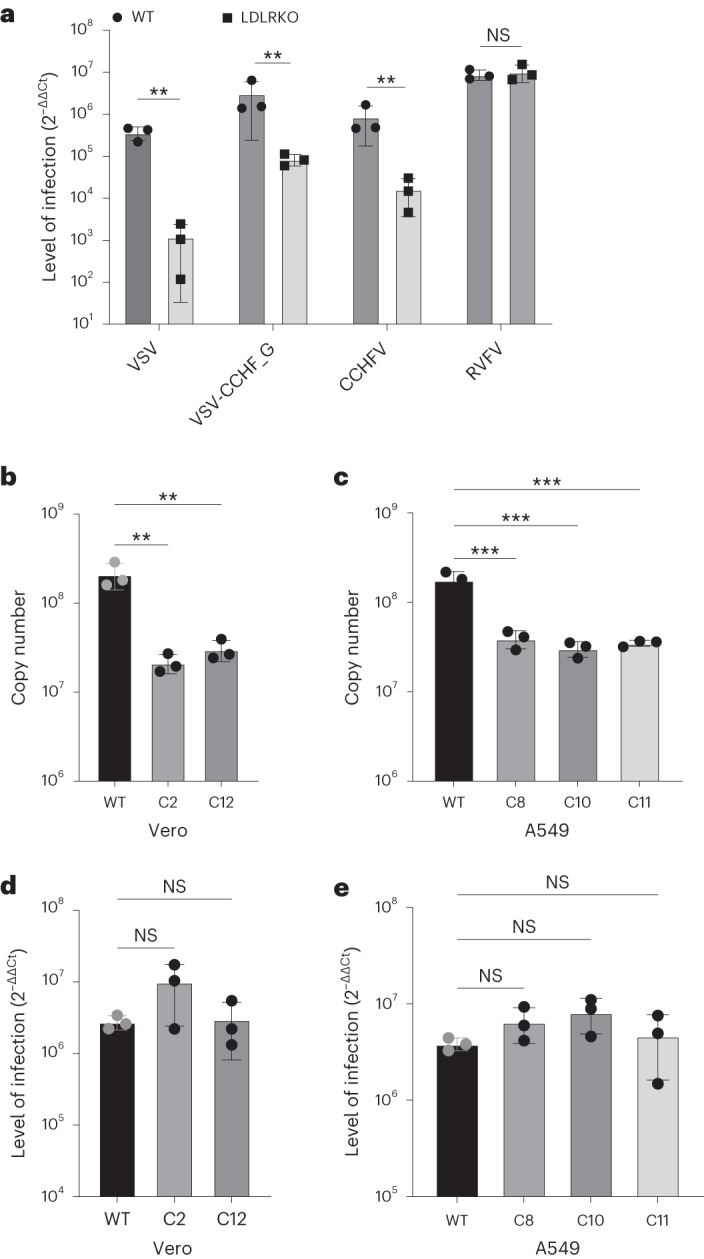
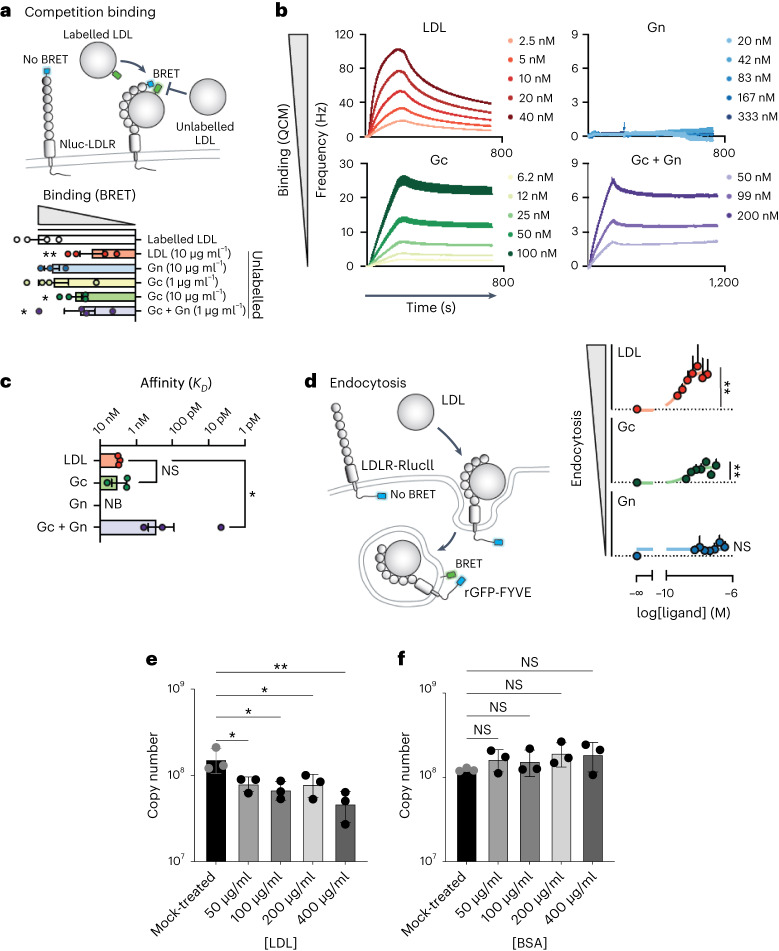
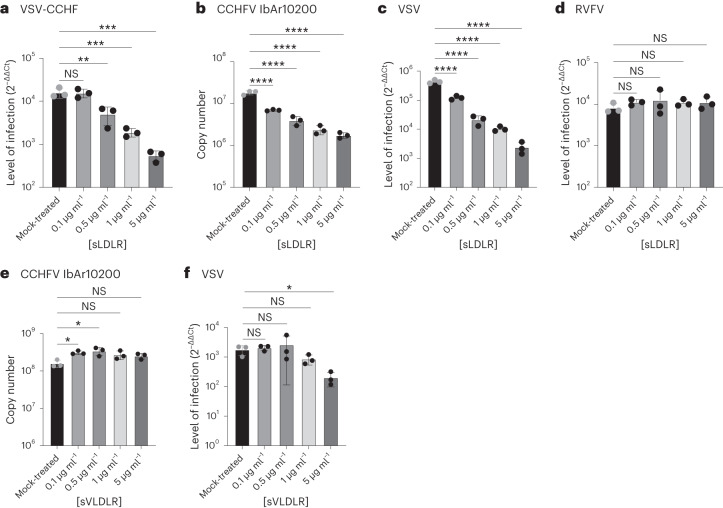
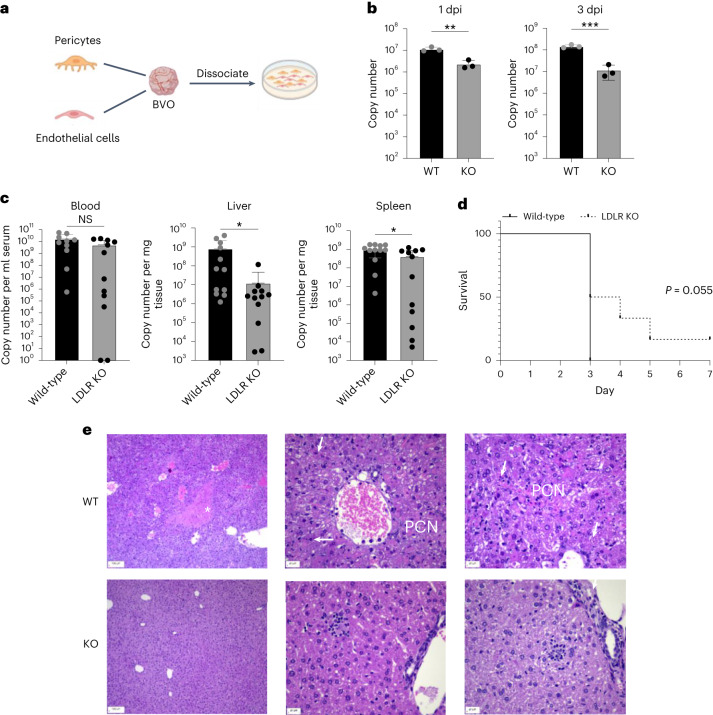
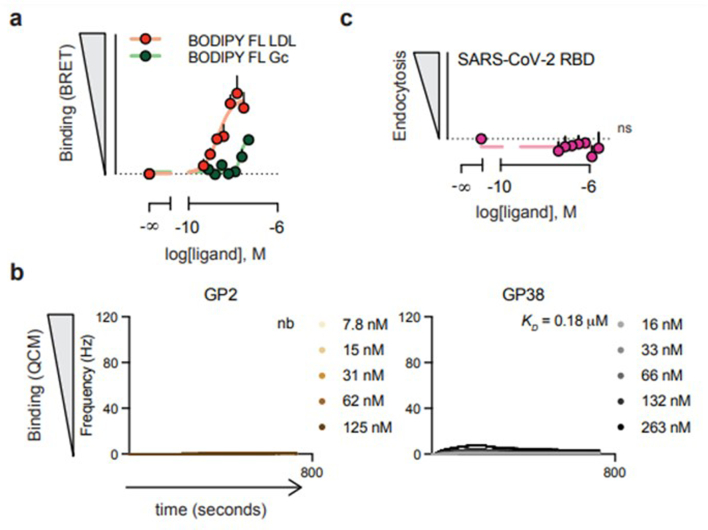
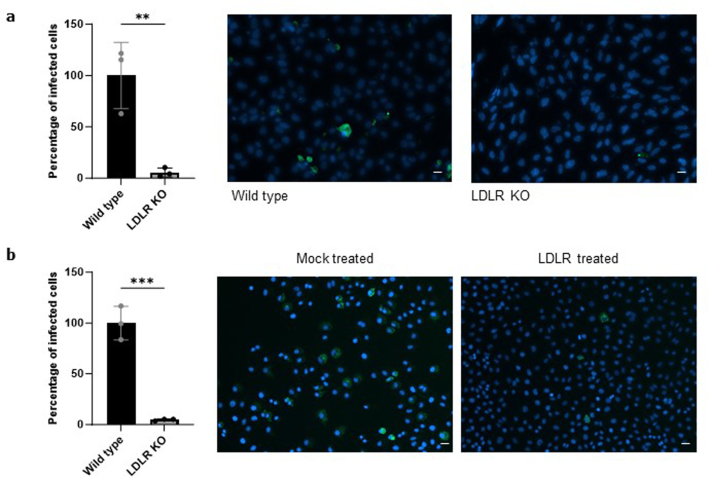
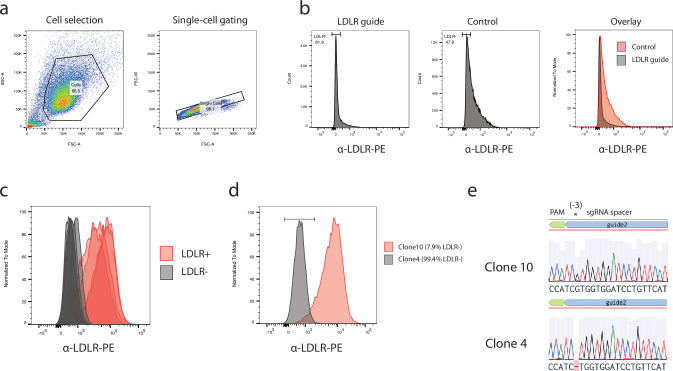
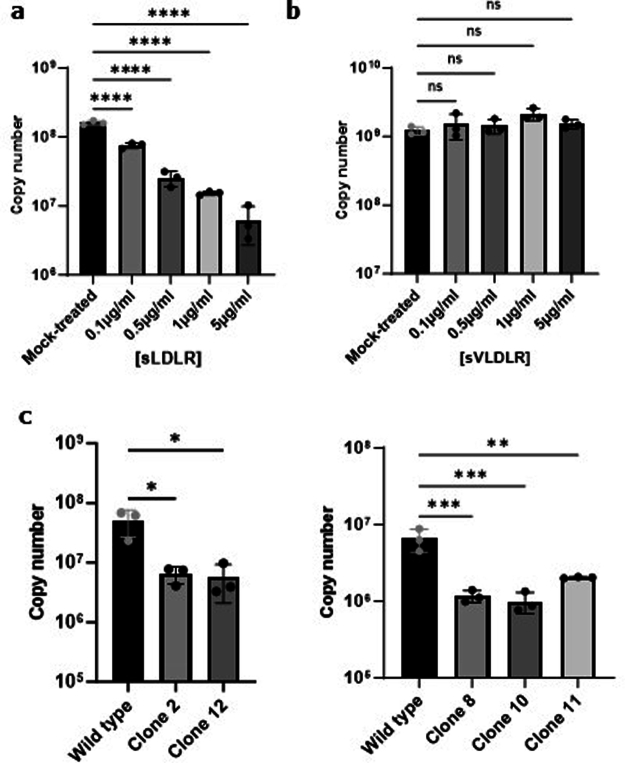
References
-
- Cases of Crimean–Congo Haemorrhagic Fever in the EU/EEA, 2013–Present (ECDC, 2024); https://www.ecdc.europa.eu/en/crimean-congo-haemorrhagic-fever/surveilla...
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
