

Long-duration environmental biosensing by recording analyte detection in DNA using recombinase memory
- PMID: 38551351
- PMCID: PMC11022584
- DOI: 10.1128/aem.02363-23
Long-duration environmental biosensing by recording analyte detection in DNA using recombinase memory
Abstract
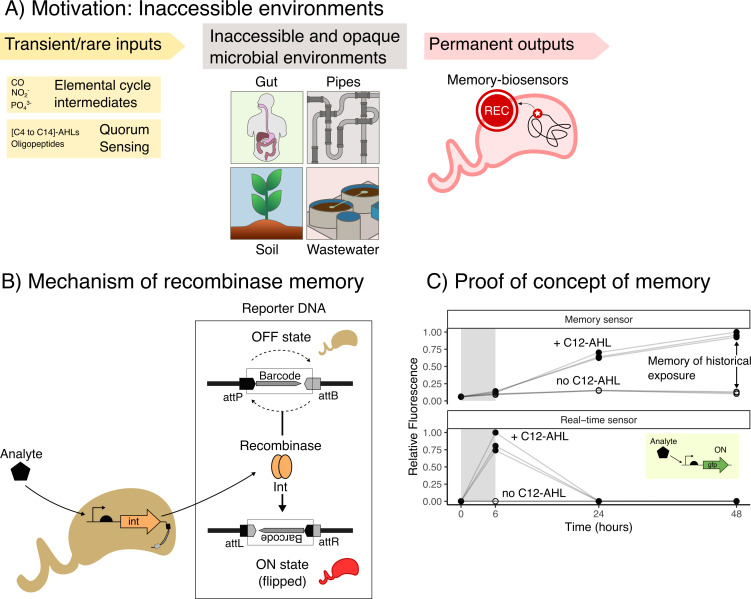
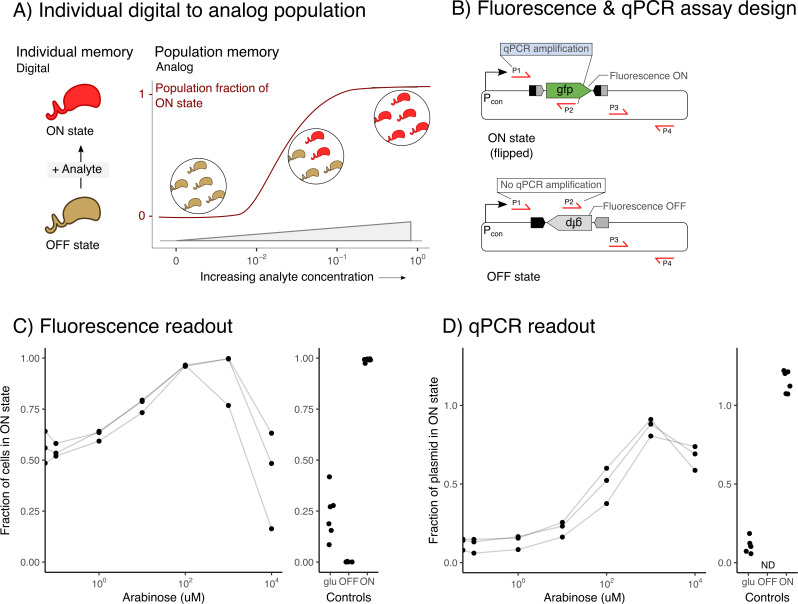
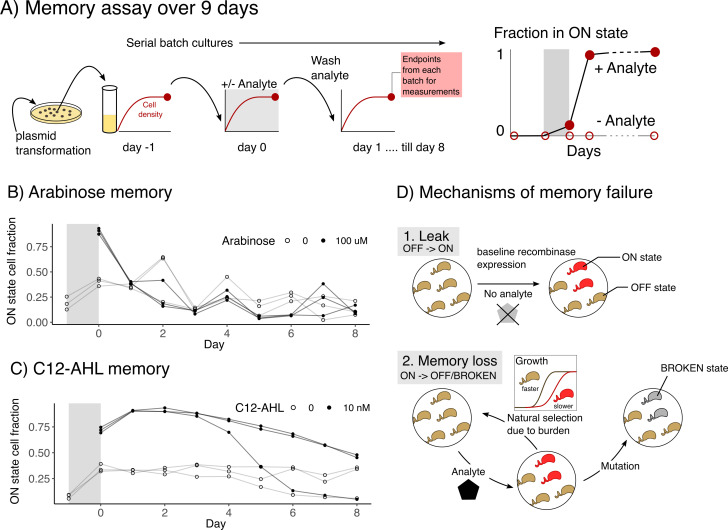
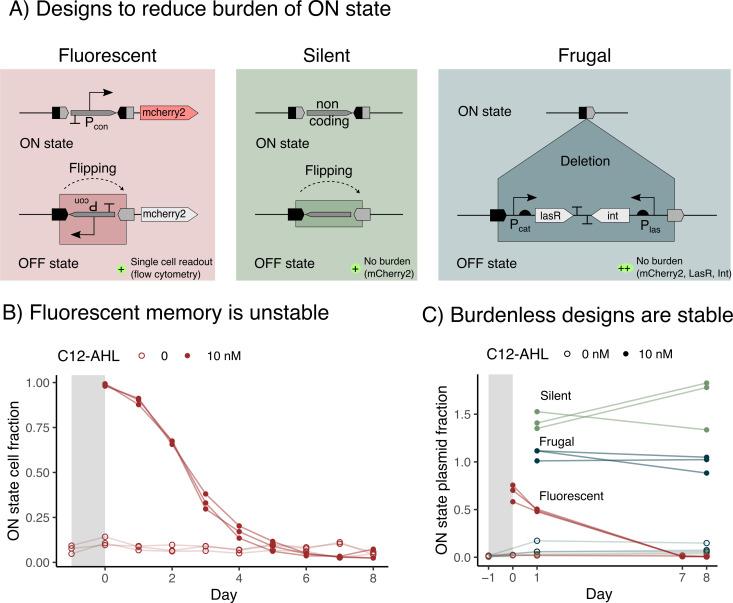
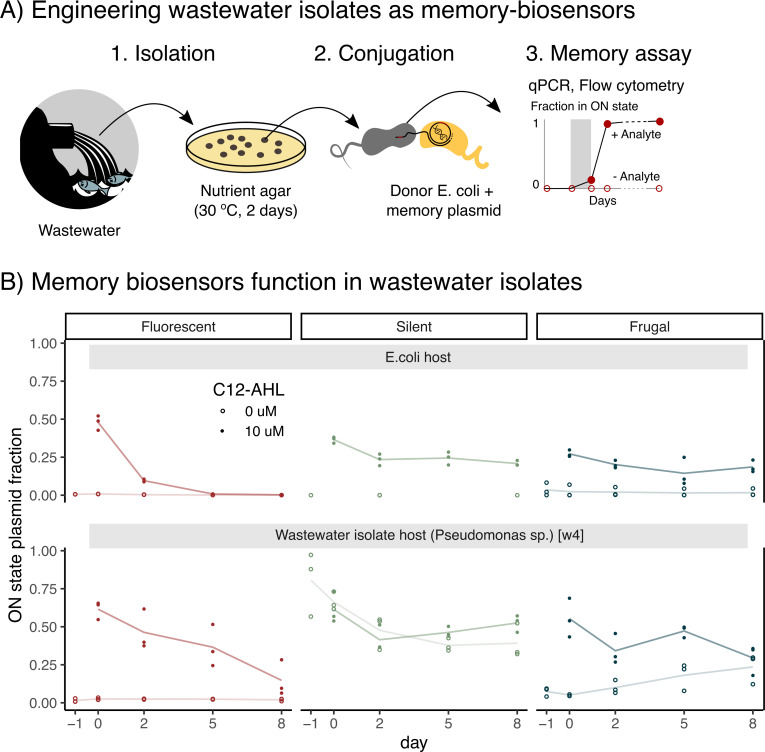
Microbial biosensors that convert environmental information into real-time visual outputs are limited in their sensing abilities in complex environments, such as soil and wastewater, due to optical inaccessibility. Biosensors that could record transient exposure to analytes within a large time window for later retrieval represent a promising approach to solve the accessibility problem. Here, we test the performance of recombinase-memory biosensors that sense a sugar (arabinose) and a microbial communication molecule (3-oxo-C12-L-homoserine lactone) over 8 days (~70 generations) following analyte exposure. These biosensors sense the analyte and trigger the expression of a recombinase enzyme which flips a segment of DNA, creating a genetic memory, and initiates fluorescent protein expression. The initial designs failed over time due to unintended DNA flipping in the absence of the analyte and loss of the flipped state after exposure to the analyte. Biosensor performance was improved by decreasing recombinase expression, removing the fluorescent protein output, and using quantitative PCR to read out stored information. Application of memory biosensors in wastewater isolates achieved memory of analyte exposure in an uncharacterized Pseudomonas isolate. By returning these engineered isolates to their native environments, recombinase-memory systems are expected to enable longer duration and in situ investigation of microbial signaling, cross-feeding, community shifts, and gene transfer beyond the reach of traditional environmental biosensors.IMPORTANCEMicrobes mediate ecological processes over timescales that can far exceed the half-lives of transient metabolites and signals that drive their collective behaviors. We investigated strategies for engineering microbes to stably record their transient exposure to a chemical over many generations through DNA rearrangements. We identify genetic architectures that improve memory biosensor performance and characterize these in wastewater isolates. Memory biosensors are expected to be useful for monitoring cell-cell signals in biofilms, detecting transient exposure to chemical pollutants, and observing microbial cross-feeding through short-lived metabolites within cryptic methane, nitrogen, and sulfur cycling processes. They will also enable in situ studies of microbial responses to ephemeral environmental changes, or other ecological processes that are currently challenging to monitor non-destructively using real-time biosensors and analytical instruments.
Keywords: biosensor; genetic memory; integrase; quorum sensing; recombinase; synthetic biology; wastewater.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Translating New Synthetic Biology Advances for Biosensing Into the Earth and Environmental Sciences.Front Microbiol. 2021 Feb 4;11:618373. doi: 10.3389/fmicb.2020.618373. eCollection 2020. Front Microbiol. 2021. PMID: 33633695 Free PMC article. Review.
-
Riboswitch-Based Reversible Dual Color Sensor.ACS Synth Biol. 2017 May 19;6(5):766-781. doi: 10.1021/acssynbio.6b00199. Epub 2017 Feb 9. ACS Synth Biol. 2017. PMID: 28121427
-
A Whole-Cell Biosensor for Detection of 2,4-Diacetylphloroglucinol (DAPG)-Producing Bacteria from Grassland Soil.Appl Environ Microbiol. 2021 Jan 15;87(3):e01400-20. doi: 10.1128/AEM.01400-20. Print 2021 Jan 15. Appl Environ Microbiol. 2021. PMID: 33218996 Free PMC article.
-
Recent advances in the analytical strategies of microbial biosensor for detection of pollutants.Chemosphere. 2022 Nov;306:135515. doi: 10.1016/j.chemosphere.2022.135515. Epub 2022 Jun 27. Chemosphere. 2022. PMID: 35772520 Review.
-
Perspective on the development of synthetic microbial community (SynCom) biosensors.Trends Biotechnol. 2023 Oct;41(10):1227-1236. doi: 10.1016/j.tibtech.2023.04.007. Epub 2023 May 13. Trends Biotechnol. 2023. PMID: 37183053 Review.
Cited by
-
Toward DNA-Based Recording of Biological Processes.Int J Mol Sci. 2024 Aug 26;25(17):9233. doi: 10.3390/ijms25179233. Int J Mol Sci. 2024. PMID: 39273181 Free PMC article. Review.
-
An autonomous microbial sensor enables long-term detection of TNT explosive in natural soil.Nat Commun. 2024 Dec 2;15(1):10471. doi: 10.1038/s41467-024-54866-y. Nat Commun. 2024. PMID: 39622841 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
