

Mastigoneme structure reveals insights into the O-linked glycosylation code of native hydroxyproline-rich helices
- PMID: 38552624
- PMCID: PMC11015965
- DOI: 10.1016/j.cell.2024.03.005
Mastigoneme structure reveals insights into the O-linked glycosylation code of native hydroxyproline-rich helices
Abstract
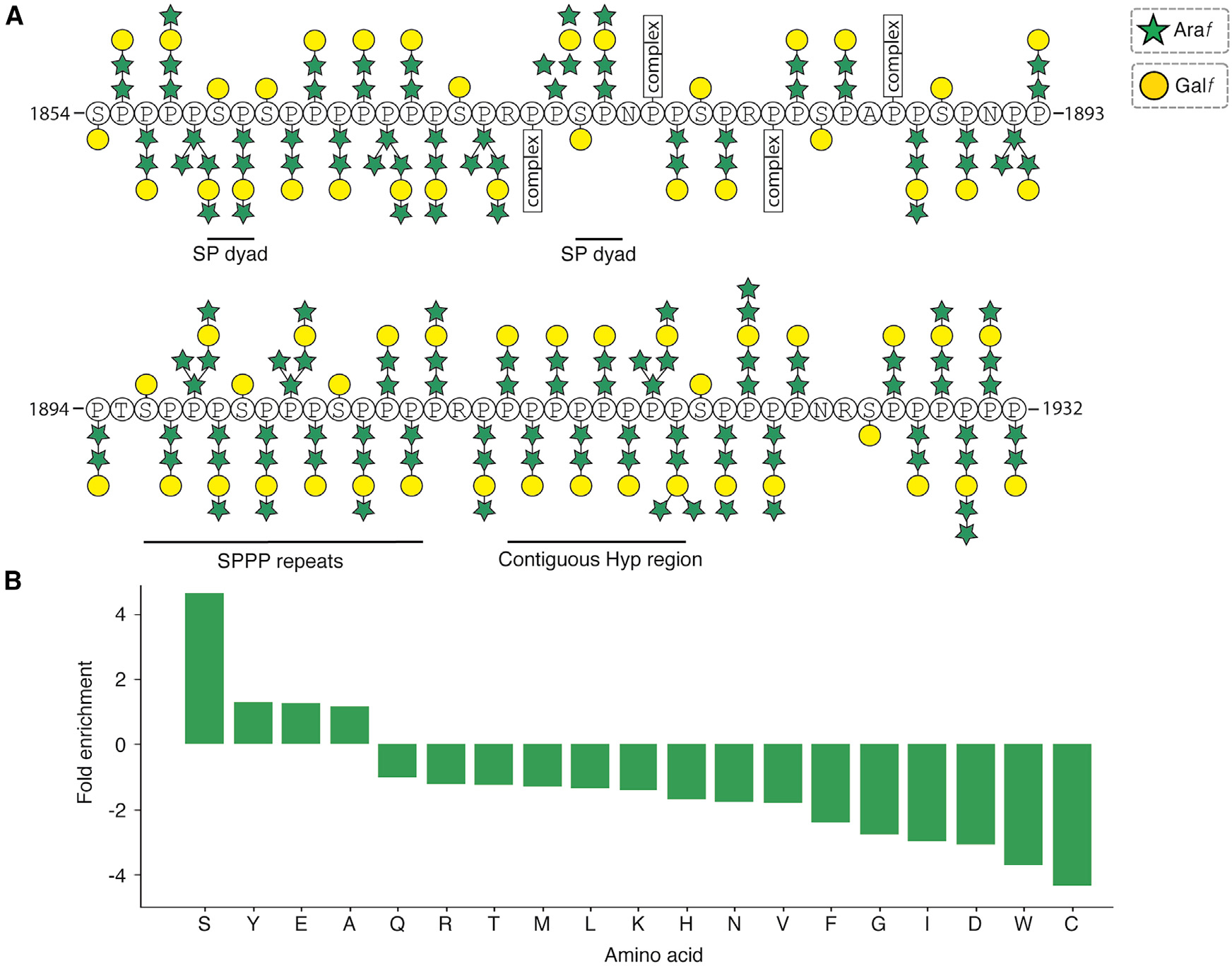
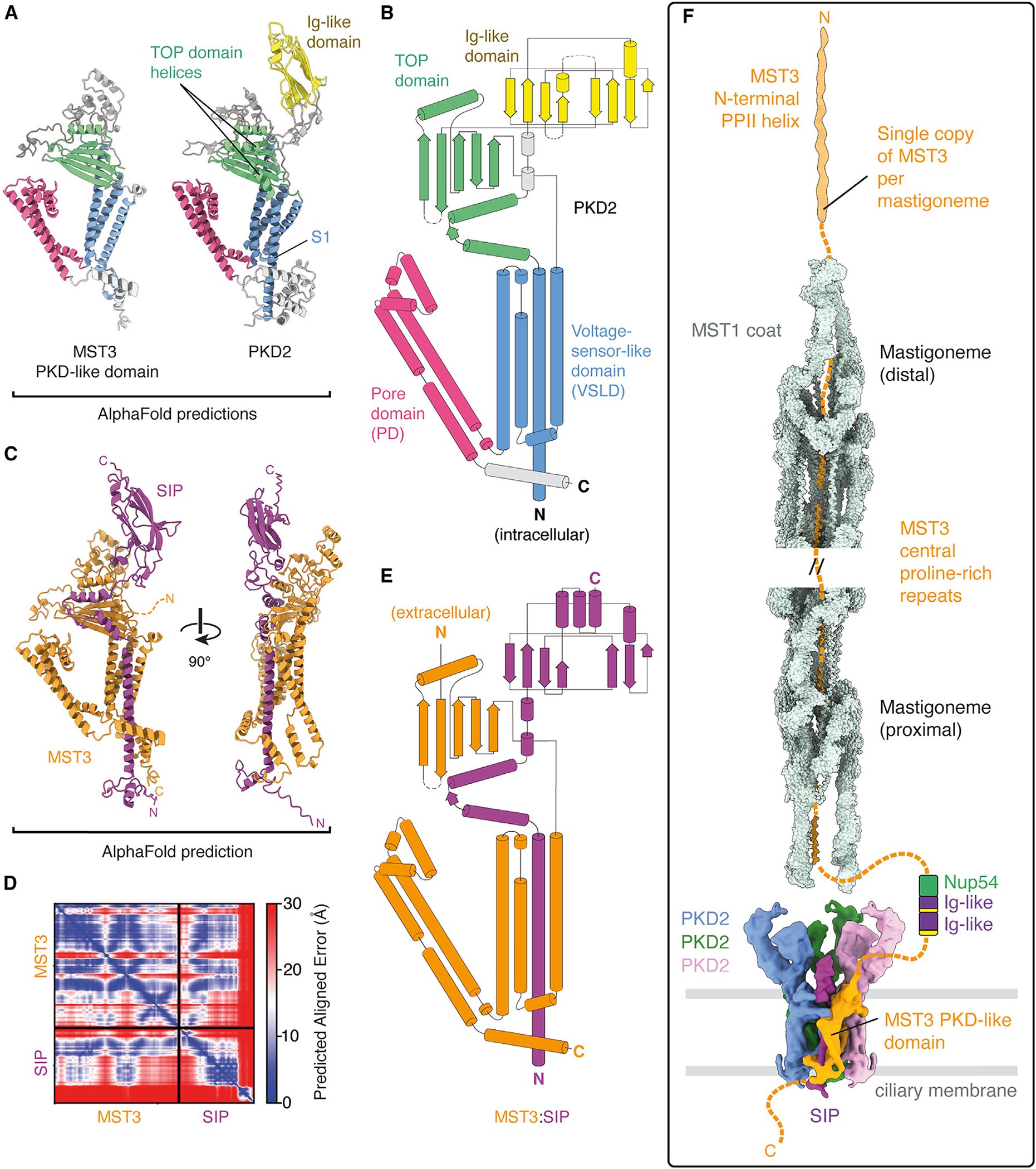
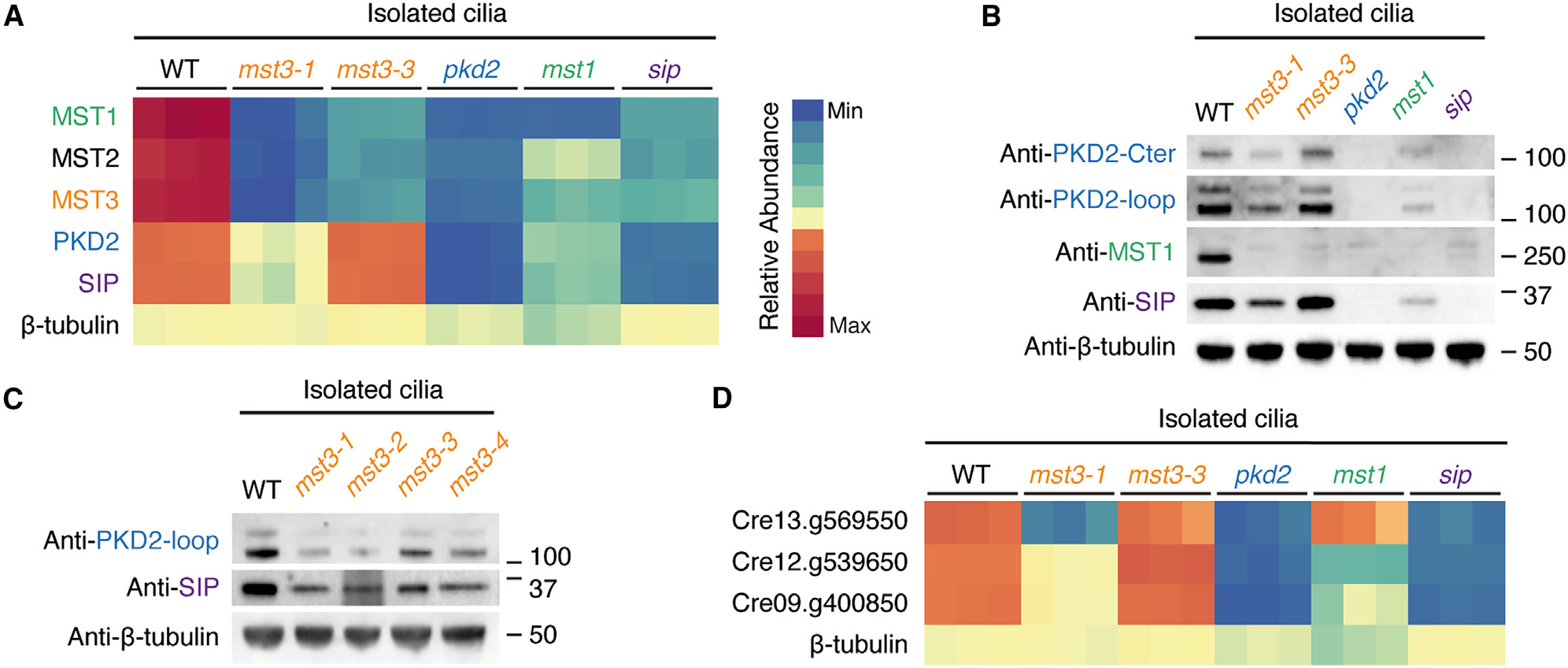
Hydroxyproline-rich glycoproteins (HRGPs) are a ubiquitous class of protein in the extracellular matrices and cell walls of plants and algae, yet little is known of their native structures or interactions. Here, we used electron cryomicroscopy (cryo-EM) to determine the structure of the hydroxyproline-rich mastigoneme, an extracellular filament isolated from the cilia of the alga Chlamydomonas reinhardtii. The structure demonstrates that mastigonemes are formed from two HRGPs (a filament of MST1 wrapped around a single copy of MST3) that both have hyperglycosylated poly(hydroxyproline) helices. Within the helices, O-linked glycosylation of the hydroxyproline residues and O-galactosylation of interspersed serine residues create a carbohydrate casing. Analysis of the associated glycans reveals how the pattern of hydroxyproline repetition determines the type and extent of glycosylation. MST3 possesses a PKD2-like transmembrane domain that forms a heteromeric polycystin-like cation channel with PKD2 and SIP, explaining how mastigonemes are tethered to ciliary membranes.
Keywords: O-linked glycosylation; cilia; cryo-EM; hydroxyproline; ion channels; mass spectrometry; mastigoneme; polycystic kidney disease; post-translational modifications.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures




Similar articles
-
Structure-guided discovery of protein and glycan components in native mastigonemes.Cell. 2024 Mar 28;187(7):1733-1744.e12. doi: 10.1016/j.cell.2024.02.037. Cell. 2024. PMID: 38552612
-
The Small Interactor of PKD2 protein promotes the assembly and ciliary entry of the Chlamydomonas PKD2-mastigoneme complexes.J Cell Sci. 2024 Jan 1;137(1):jcs261497. doi: 10.1242/jcs.261497. Epub 2024 Jan 12. J Cell Sci. 2024. PMID: 38063216 Free PMC article.
-
Chlamydomonas PKD2 organizes mastigonemes, hair-like glycoprotein polymers on cilia.J Cell Biol. 2020 Jun 1;219(6):e202001122. doi: 10.1083/jcb.202001122. J Cell Biol. 2020. PMID: 32348466 Free PMC article.
-
An update on post-translational modifications of hydroxyproline-rich glycoproteins: toward a model highlighting their contribution to plant cell wall architecture.Front Plant Sci. 2014 Aug 14;5:395. doi: 10.3389/fpls.2014.00395. eCollection 2014. Front Plant Sci. 2014. PMID: 25177325 Free PMC article. Review.
-
Ciliary mechanosensation - roles of polycystins and mastigonemes.J Cell Sci. 2023 Feb 1;136(3):jcs260565. doi: 10.1242/jcs.260565. Epub 2023 Feb 8. J Cell Sci. 2023. PMID: 36752106 Review.
Cited by
-
Unwrapping the Ciliary Coat: High-Resolution Structure and Function of the Ciliary Glycocalyx.Adv Sci (Weinh). 2025 Apr;12(16):e2413355. doi: 10.1002/advs.202413355. Epub 2025 Mar 5. Adv Sci (Weinh). 2025. PMID: 40041987 Free PMC article.
-
BMP4 regulates asymmetric Pkd2 distribution in mouse nodal immotile cilia and ciliary mechanosensing required for left-right determination.Dev Dyn. 2025 Aug;254(8):965-978. doi: 10.1002/dvdy.727. Epub 2024 Jul 10. Dev Dyn. 2025. PMID: 38984461 Free PMC article.
References
-
- Johnson KL, Cassin AM, Lonsdale A, Wong GK-S, Soltis DE, Miles NW, Melkonian M, Melkonian B, Deyholos MK, Leebens-Mack J, et al. (2017). Insights into the Evolution of Hydroxyproline-Rich Glycoproteins from 1000 Plant Transcriptomes. Plant Physiol. 174, 904–921. 10.1104/pp.17.00295. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
