

Menke-Hennekam syndrome; delineation of domain-specific subtypes with distinct clinical and DNA methylation profiles
- PMID: 38553851
- PMCID: PMC11040166
- DOI: 10.1016/j.xhgg.2024.100287
Menke-Hennekam syndrome; delineation of domain-specific subtypes with distinct clinical and DNA methylation profiles
Erratum in
-
Menke-Hennekam syndrome; delineation of domain-specific subtypes with distinct clinical and DNA methylation profiles.HGG Adv. 2024 Oct 10;5(4):100337. doi: 10.1016/j.xhgg.2024.100337. Epub 2024 Sep 21. HGG Adv. 2024. PMID: 39306848 Free PMC article. No abstract available.
Abstract
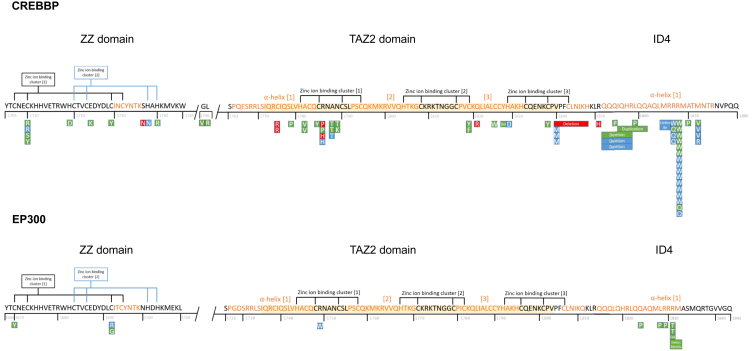
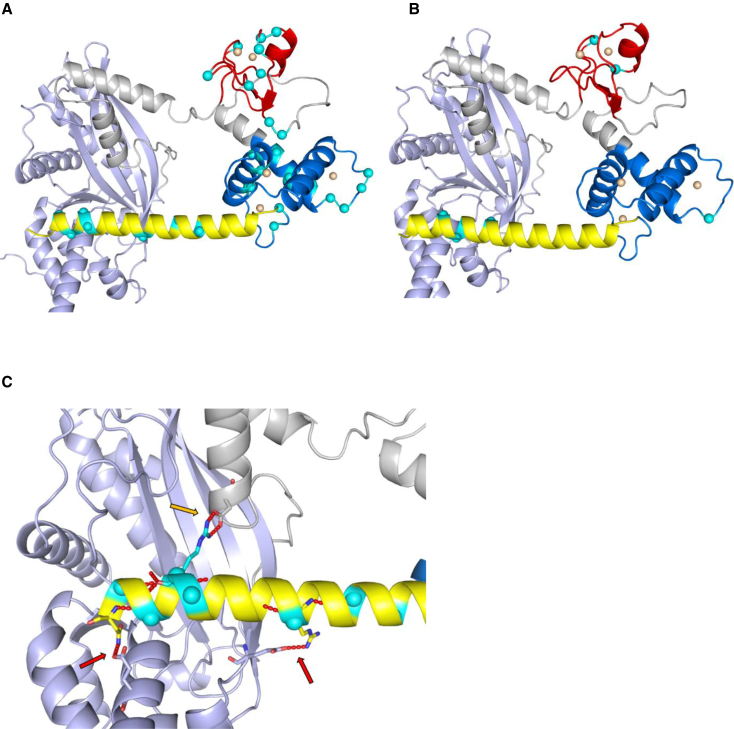
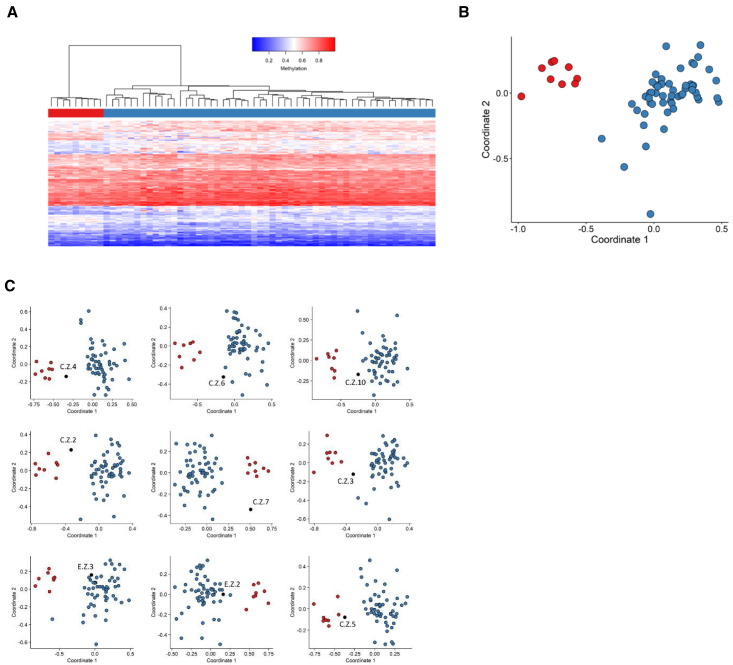
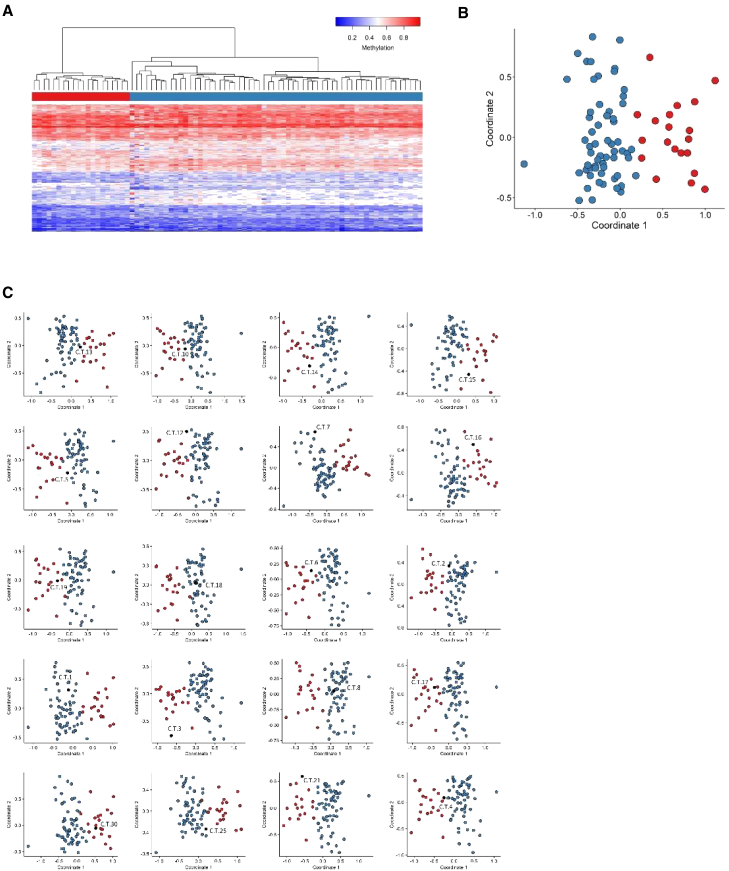
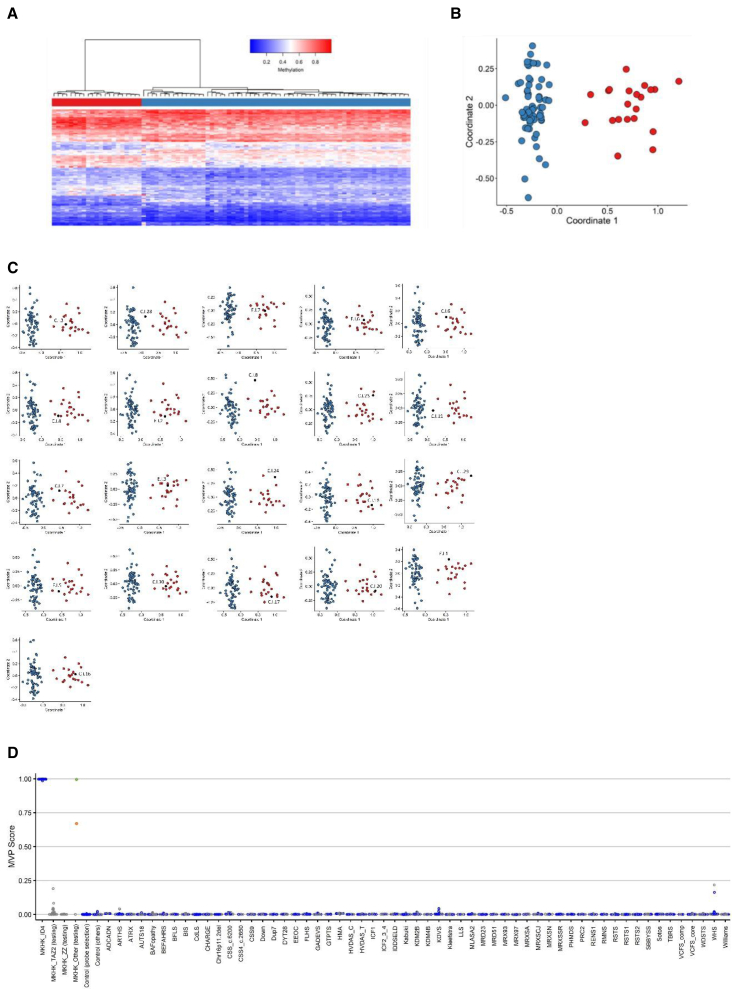
CREB-binding protein (CBP, encoded by CREBBP) and its paralog E1A-associated protein (p300, encoded by EP300) are involved in histone acetylation and transcriptional regulation. Variants that produce a null allele or disrupt the catalytic domain of either protein cause Rubinstein-Taybi syndrome (RSTS), while pathogenic missense and in-frame indel variants in parts of exons 30 and 31 cause phenotypes recently described as Menke-Hennekam syndrome (MKHK). To distinguish MKHK subtypes and define their characteristics, molecular and extended clinical data on 82 individuals (54 unpublished) with variants affecting CBP (n = 71) or p300 (n = 11) (NP_004371.2 residues 1,705-1,875 and NP_001420.2 residues 1,668-1,833, respectively) were summarized. Additionally, genome-wide DNA methylation profiles were assessed in DNA extracted from whole peripheral blood from 54 individuals. Most variants clustered closely around the zinc-binding residues of two zinc-finger domains (ZZ and TAZ2) and within the first α helix of the fourth intrinsically disordered linker (ID4) of CBP/p300. Domain-specific methylation profiles were discerned for the ZZ domain in CBP/p300 (found in nine out of 10 tested individuals) and TAZ2 domain in CBP (in 14 out of 20), while a domain-specific diagnostic episignature was refined for the ID4 domain in CBP/p300 (in 21 out of 21). Phenotypes including intellectual disability of varying degree and distinct physical features were defined for each of the regions. These findings demonstrate existence of at least three MKHK subtypes, which are domain specific (MKHK-ZZ, MKHK-TAZ2, and MKHK-ID4) rather than gene specific (CREBBP/EP300). DNA methylation episignatures enable stratification of molecular pathophysiologic entities within a gene or across a family of paralogous genes.
Keywords: CREB-binding protein; DNA methylation; E1A-associated protein p300; MKHK; Menke-Hennekam syndrome; Rubinstein-Taybi syndrome; episignatures; intellectual disability; intrinsically disordered linker; zinc-finger domain.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests B.S. is a shareholder in EpiSign Inc., a biotech firm involved in commercial application of EpiSign technology.
Figures

References
-
- Roelfsema J.H., White S.J., Ariyürek Y., Bartholdi D., Niedrist D., Papadia F., Bacino C.A., den Dunnen J.T., van Ommen G.J.B., Breuning M.H., et al. Genetic heterogeneity in Rubinstein-Taybi syndrome: mutations in both the CBP and EP300 genes cause disease. Am. J. Hum. Genet. 2005;76:572–580. doi: 10.1086/429130. - DOI - PMC - PubMed
-
- Cross E., Duncan-Flavell P.J., Howarth R.J., Hobbs J.I., Thomas N.S., Bunyan D.J. Screening of a large Rubinstein-Taybi cohort identified many novel variants and emphasizes the importance of the CREBBP histone acetyltransferase domain. Am. J. Med. Genet. 2020;182:2508–2520. doi: 10.1002/ajmg.a.61813. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
