

Microglial phagolysosome dysfunction and altered neural communication amplify phenotypic severity in Prader-Willi Syndrome with larger deletion
- PMID: 38556574
- PMCID: PMC10982101
- DOI: 10.1007/s00401-024-02714-0
Microglial phagolysosome dysfunction and altered neural communication amplify phenotypic severity in Prader-Willi Syndrome with larger deletion
Abstract
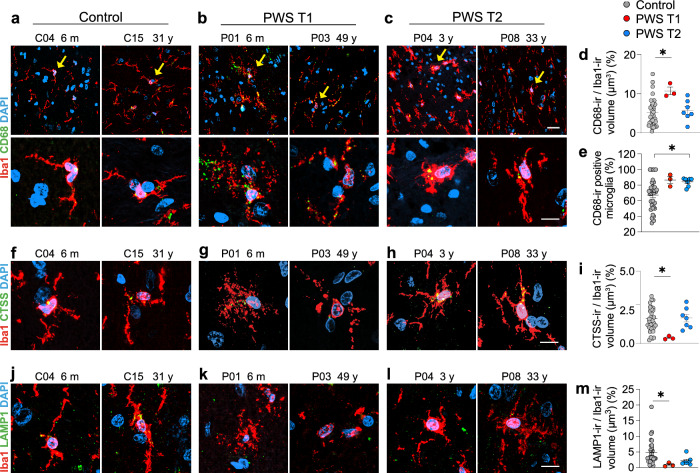
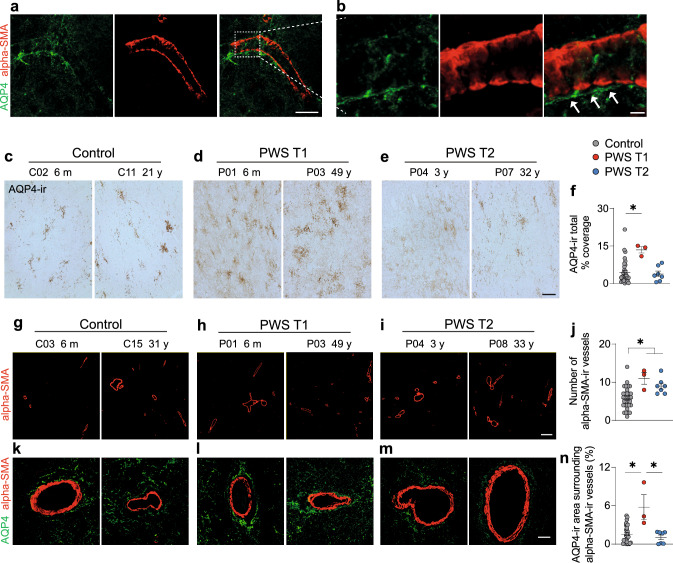
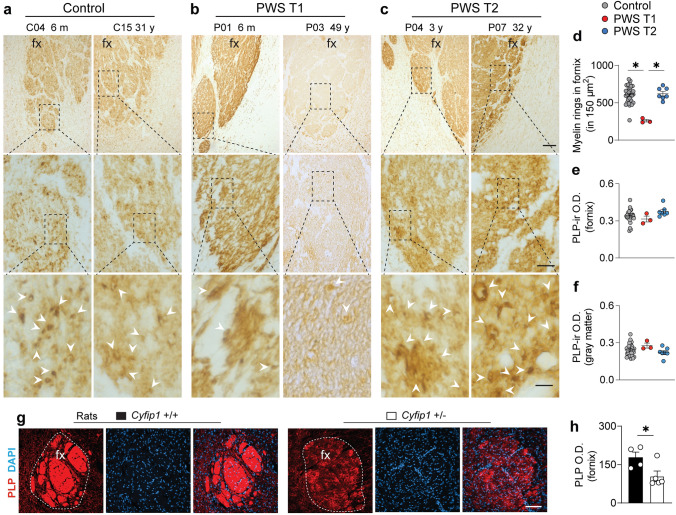
Prader-Willi Syndrome (PWS) is a rare neurodevelopmental disorder of genetic etiology, characterized by paternal deletion of genes located at chromosome 15 in 70% of cases. Two distinct genetic subtypes of PWS deletions are characterized, where type I (PWS T1) carries four extra haploinsufficient genes compared to type II (PWS T2). PWS T1 individuals display more pronounced physiological and cognitive abnormalities than PWS T2, yet the exact neuropathological mechanisms behind these differences remain unclear. Our study employed postmortem hypothalamic tissues from PWS T1 and T2 individuals, conducting transcriptomic analyses and cell-specific protein profiling in white matter, neurons, and glial cells to unravel the cellular and molecular basis of phenotypic severity in PWS sub-genotypes. In PWS T1, key pathways for cell structure, integrity, and neuronal communication are notably diminished, while glymphatic system activity is heightened compared to PWS T2. The microglial defect in PWS T1 appears to stem from gene haploinsufficiency, as global and myeloid-specific Cyfip1 haploinsufficiency in murine models demonstrated. Our findings emphasize microglial phagolysosome dysfunction and altered neural communication as crucial contributors to the severity of PWS T1's phenotype.
Keywords: Fornix; Glymphatic system; Hypothalamus; Immunosurveillance; Microglia; Myelin; Oxytocin.
© 2024. The Author(s).
Conflict of interest statement
The authors have declared that no conflict of interest exists.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
