

Anti-PD-1 chimeric antigen receptor T cells efficiently target SIV-infected CD4+ T cells in germinal centers
- PMID: 38557496
- PMCID: PMC10977982
- DOI: 10.1172/JCI169309
Anti-PD-1 chimeric antigen receptor T cells efficiently target SIV-infected CD4+ T cells in germinal centers
Abstract
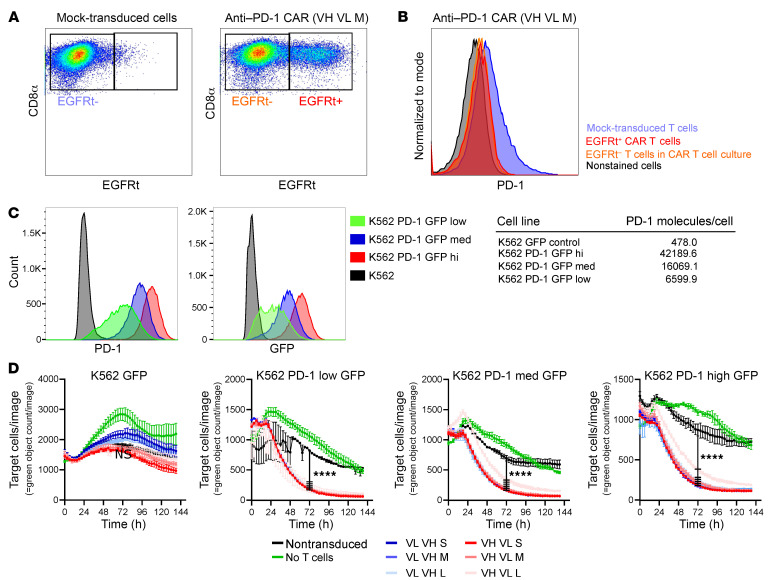
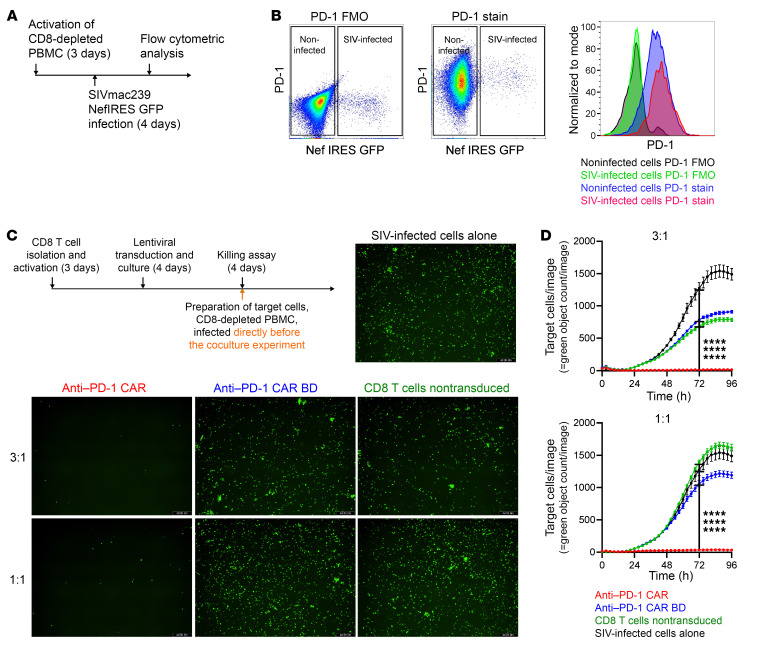
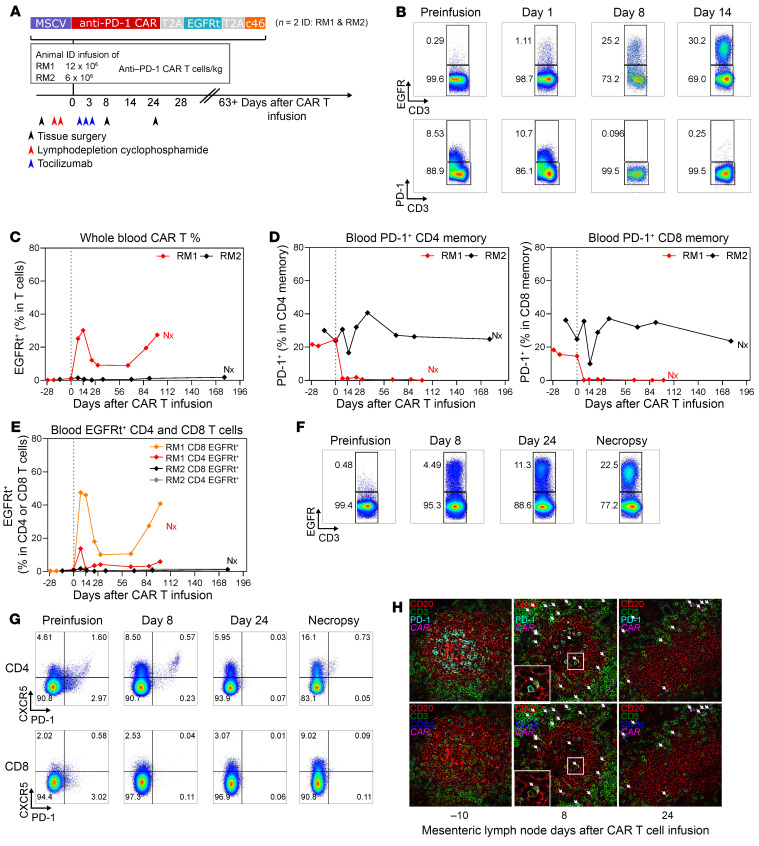
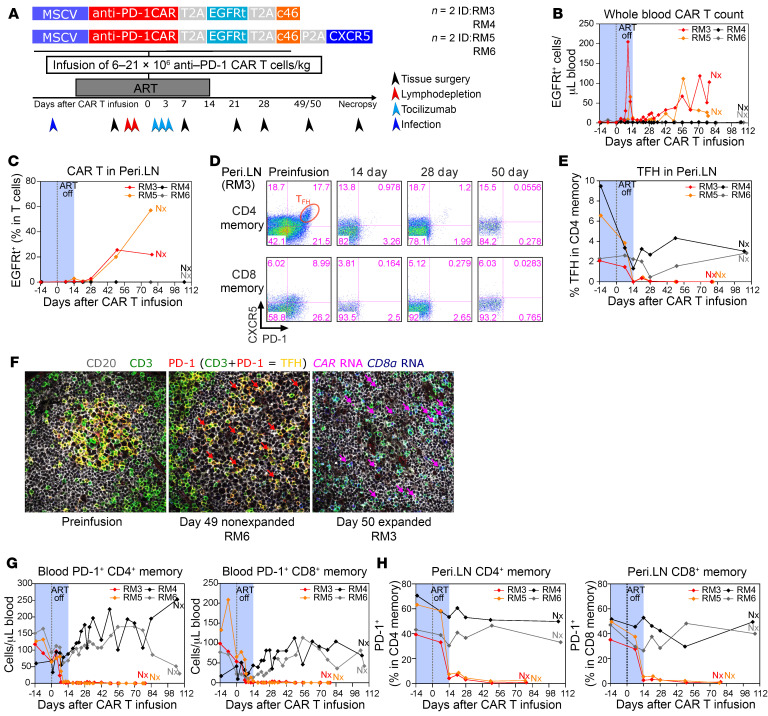
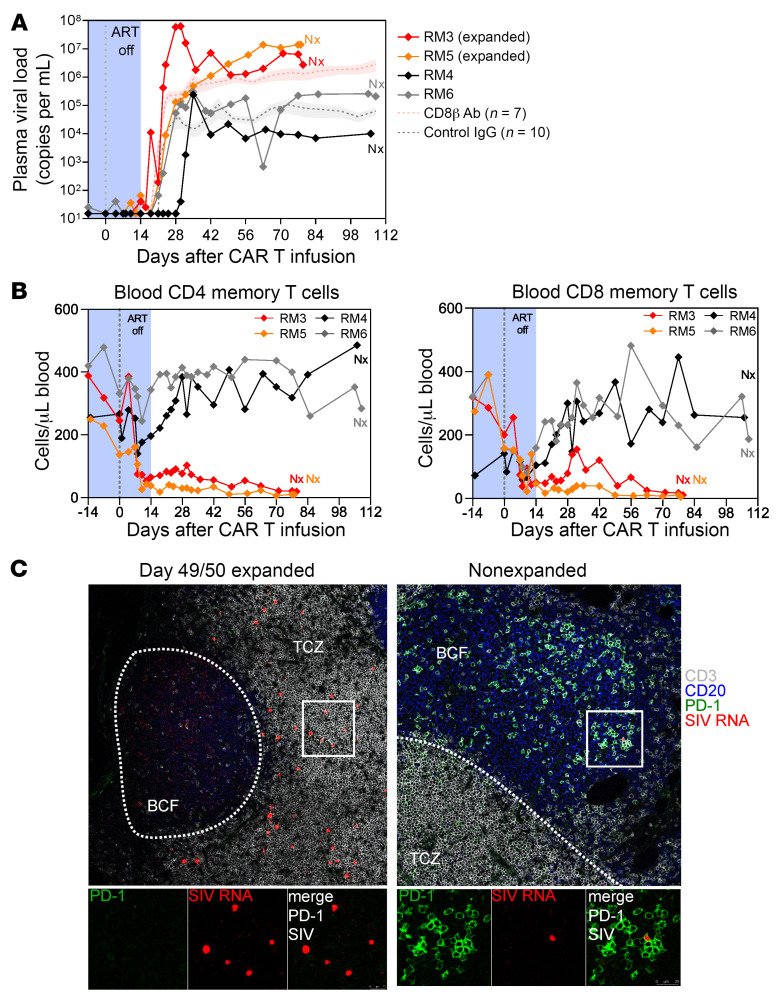
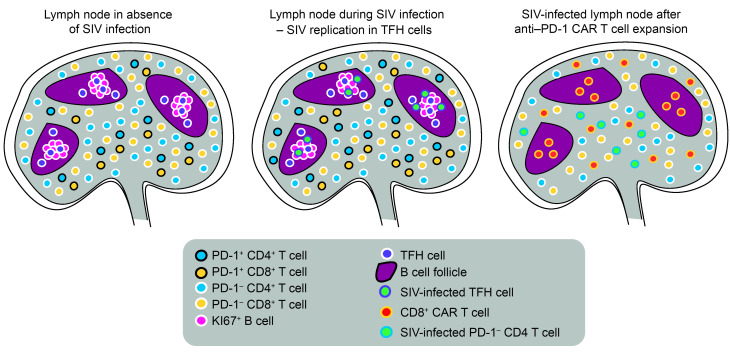
Programmed cell death protein 1 (PD-1) is an immune checkpoint marker commonly expressed on memory T cells and enriched in latently HIV-infected CD4+ T cells. We engineered an anti-PD-1 chimeric antigen receptor (CAR) to assess the impact of PD-1 depletion on viral reservoirs and rebound dynamics in SIVmac239-infected rhesus macaques (RMs). Adoptive transfer of anti-PD-1 CAR T cells was done in 2 SIV-naive and 4 SIV-infected RMs on antiretroviral therapy (ART). In 3 of 6 RMs, anti-PD-1 CAR T cells expanded and persisted for up to 100 days concomitant with the depletion of PD-1+ memory T cells in blood and tissues, including lymph node CD4+ follicular helper T (TFH) cells. Loss of TFH cells was associated with depletion of detectable SIV RNA from the germinal center (GC). However, following CAR T infusion and ART interruption, there was a marked increase in SIV replication in extrafollicular portions of lymph nodes, a 2-log higher plasma viremia relative to controls, and accelerated disease progression associated with the depletion of CD8+ memory T cells. These data indicate anti-PD-1 CAR T cells depleted PD-1+ T cells, including GC TFH cells, and eradicated SIV from this immunological sanctuary.
Keywords: AIDS/HIV; Gene therapy; Immunotherapy; T cells; Therapeutics.
Conflict of interest statement
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
