

Landscape of brain myeloid cell transcriptome along the spatiotemporal progression of Alzheimer's disease reveals distinct sequential responses to Aβ and tau
- PMID: 38557897
- PMCID: PMC10984903
- DOI: 10.1007/s00401-024-02704-2
Landscape of brain myeloid cell transcriptome along the spatiotemporal progression of Alzheimer's disease reveals distinct sequential responses to Aβ and tau
Abstract
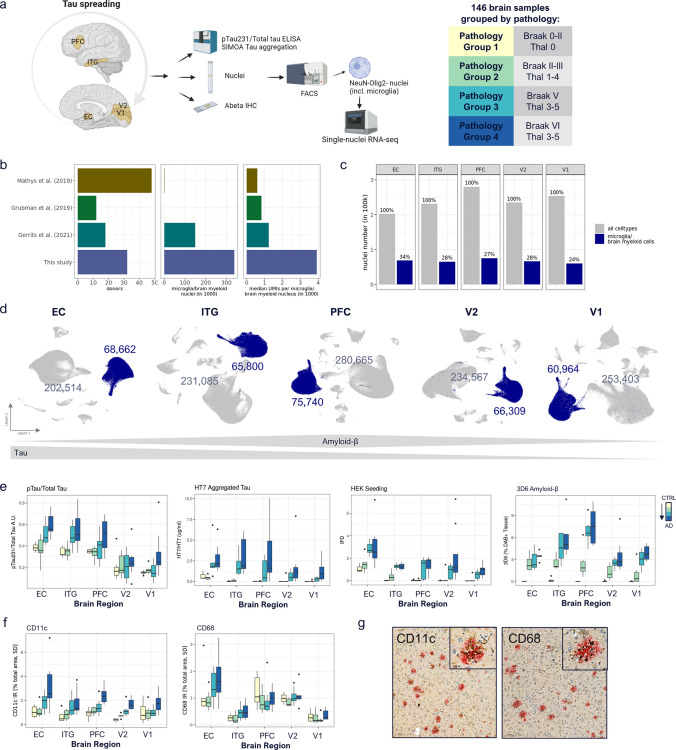
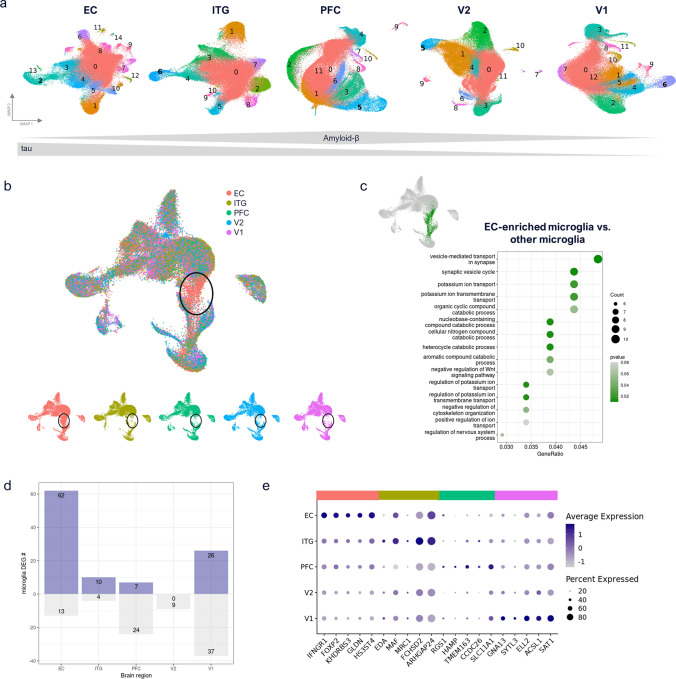
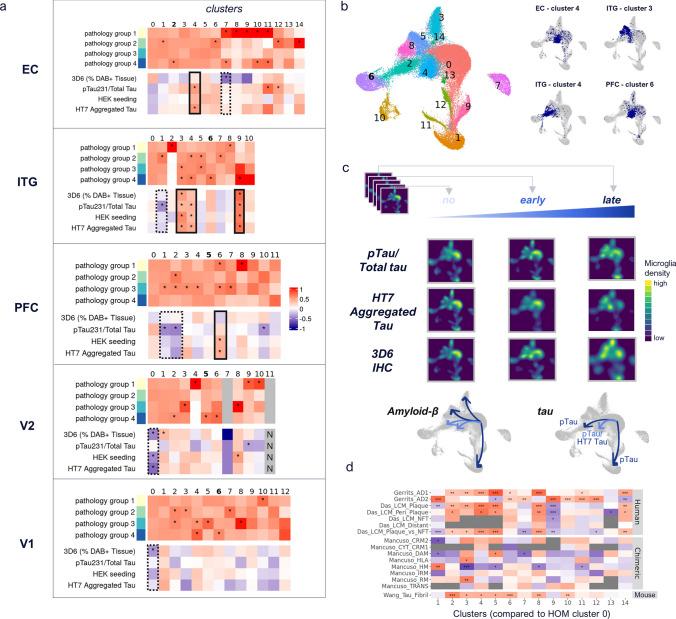
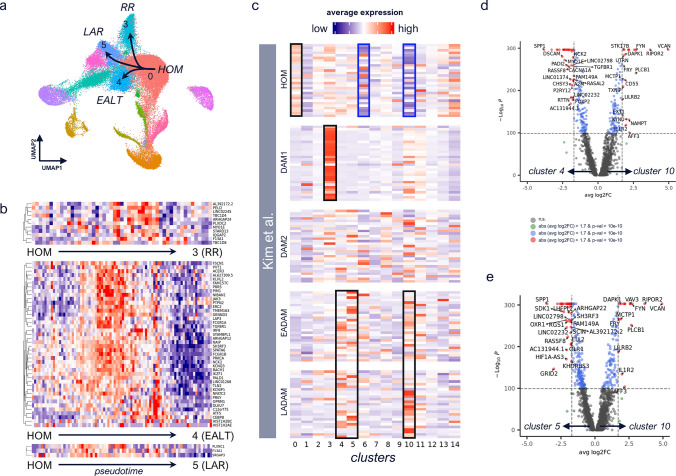
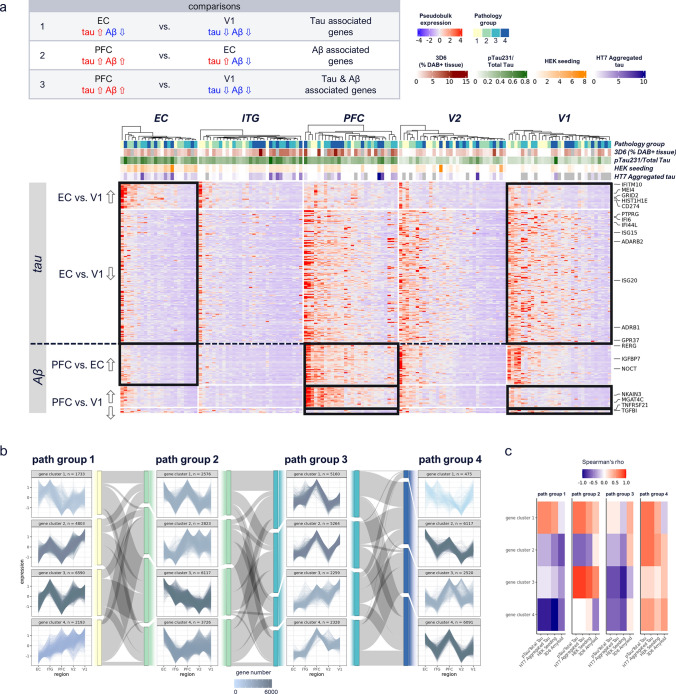
Human microglia are critically involved in Alzheimer's disease (AD) progression, as shown by genetic and molecular studies. However, their role in tau pathology progression in human brain has not been well described. Here, we characterized 32 human donors along progression of AD pathology, both in time-from early to late pathology-and in space-from entorhinal cortex (EC), inferior temporal gyrus (ITG), prefrontal cortex (PFC) to visual cortex (V2 and V1)-with biochemistry, immunohistochemistry, and single nuclei-RNA-sequencing, profiling a total of 337,512 brain myeloid cells, including microglia. While the majority of microglia are similar across brain regions, we identified a specific subset unique to EC which may contribute to the early tau pathology present in this region. We calculated conversion of microglia subtypes to diseased states and compared conversion patterns to those from AD animal models. Targeting genes implicated in this conversion, or their upstream/downstream pathways, could halt gene programs initiated by early tau progression. We used expression patterns of early tau progression to identify genes whose expression is reversed along spreading of spatial tau pathology (EC > ITG > PFC > V2 > V1) and identified their potential involvement in microglia subtype conversion to a diseased state. This study provides a data resource that builds on our knowledge of myeloid cell contribution to AD by defining the heterogeneity of microglia and brain macrophages during both temporal and regional pathology aspects of AD progression at an unprecedented resolution.
Keywords: Alzheimer’s disease; Microglia; Myeloid cells; Single-nucleus RNA-sequencing.
© 2024. The Author(s).
Conflict of interest statement
AW, MEW, SL, CW, EM, NR, KY, GL, FL, CK, YG, MPK, TD, EHK, XL, JSR, RVT, and KB are employees of AbbVie. AA, TP, JT, and TK were AbbVie employees at the time of the study. The design, study conduct, and financial support for this research were provided by AbbVie. AbbVie participated in the interpretation of data, review, and approval of the publication. BTH has received research funding from AbbVie as part of a collaboration agreement with The General Hospital Corporation, d/b/a Massachusetts General Hospital. BTH has a family member who works at Novartis and owns stock in Novartis; he serves on the SAB of Dewpoint and owns stock. He serves on a scientific advisory board or is a consultant for AbbVie, Aprinoia, Avrobio, Biogen, BMS Cell Signaling, Genentech, Novartis, Seer, Takeda, the US Dept of Justice, Vigil, and Voyager. His laboratory is supported by sponsored research agreements with AbbVie, and research grants from the National Institutes of Health, Cure Alzheimer’s Fund, Tau Consortium, and the JPB Foundation. MPF has Sponsored Research Agreements with Biogen and Voyager Therapeutics and works on the AbbVie-Hyman Collaboration. ASP, MJ, HL, SD, and RB work on the AbbVie-Hyman Collaboration and have no other funding to disclose.
Figures
References
-
- Ahuja M, Kaidery NA, Attucks OC, McDade E, Hushpulian DM, Gaisin A, Gaisina I, Ahn YH, Nikulin S, Poloznikov A, Gazaryan I, Yamamoto M, Matsumoto M, Igarashi K, Sharma SM, Thomas B. Bach1 derepression is neuroprotective in a mouse model of Parkinson’s disease. Proc Natl Acad Sci. 2021;118:e2111643118. doi: 10.1073/pnas.2111643118. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
