

This is a preprint.
Meiosis-specific functions of kinetochore protein SPC105R required for chromosome segregation in Drosophila oocytes
- PMID: 38559067
- PMCID: PMC10980020
- DOI: 10.1101/2024.03.14.585003
Meiosis-specific functions of kinetochore protein SPC105R required for chromosome segregation in Drosophila oocytes
Update in
-
Meiosis-specific functions of kinetochore protein SPC105R required for chromosome segregation in Drosophila oocytes.Mol Biol Cell. 2024 Aug 1;35(8):ar105. doi: 10.1091/mbc.E24-02-0067. Epub 2024 Jun 12. Mol Biol Cell. 2024. PMID: 38865189 Free PMC article.
Abstract
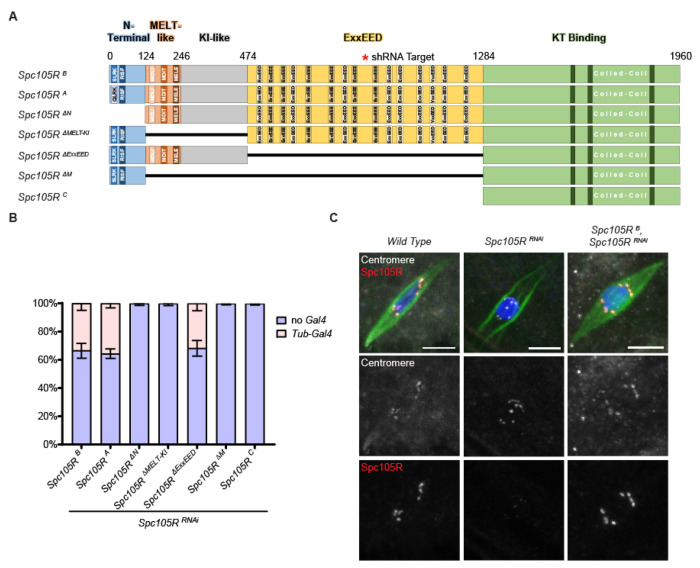
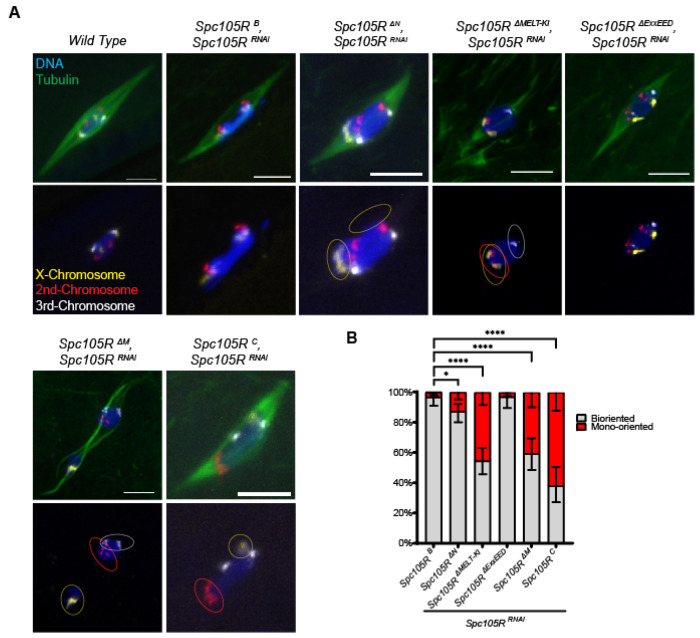
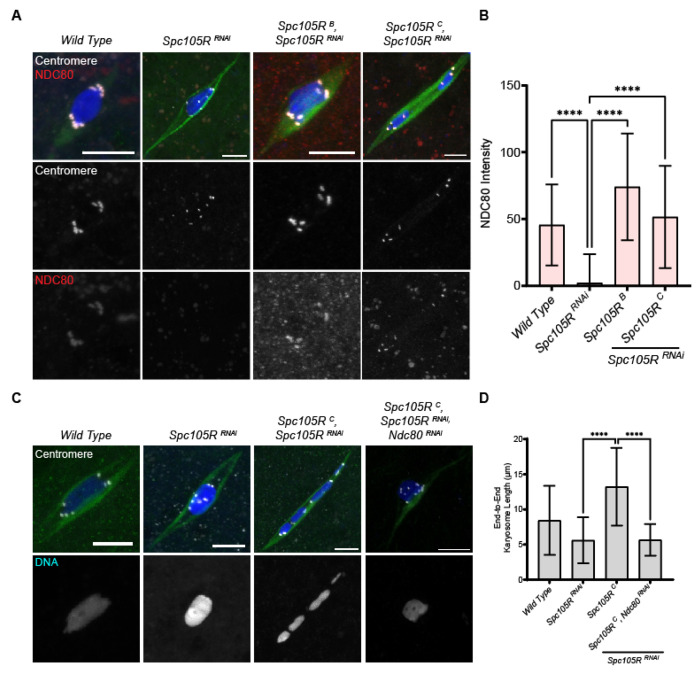
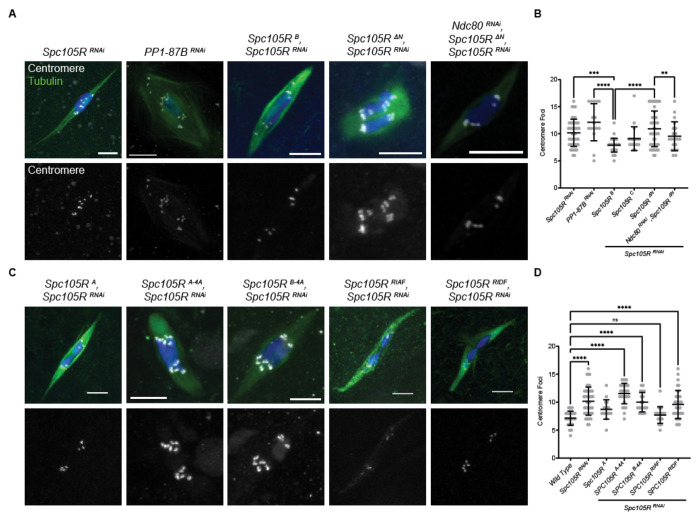
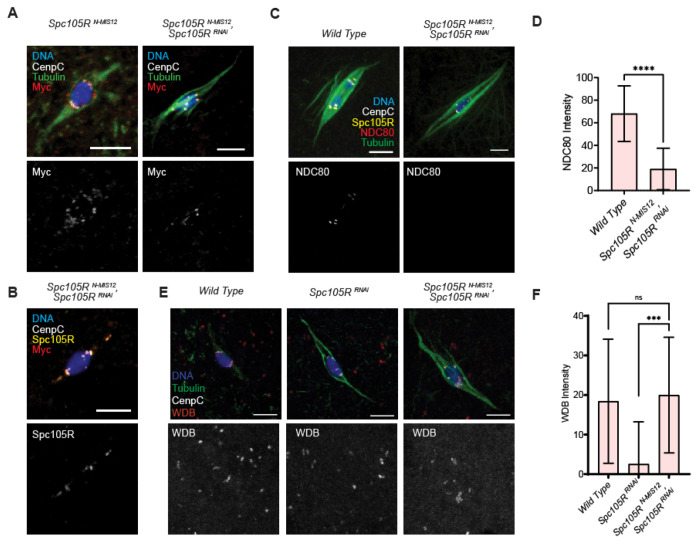
The reductional division of meiosis I requires the separation of chromosome pairs towards opposite poles. We have previously implicated the outer kinetochore protein SPC105R/KNL1 in driving meiosis I chromosome segregation through lateral attachments to microtubules and co-orientation of sister centromeres. To identify the domains of SPC105R that are critical for meiotic chromosome segregation, an RNAi-resistant gene expression system was developed. We found that SPC105R's C-terminal domain (aa 1284-1960) is necessary and sufficient for recruiting NDC80 to the kinetochore and building the outer kinetochore. Furthermore, the C-terminal domain recruits BUBR1, which in turn recruits the cohesion protection proteins MEI-S332 and PP2A. Of the remaining 1283 amino acids, we found the first 473 are most important for meiosis. The first 123 amino acids of the N-terminal half of SPC105R contain the conserved SLRK and RISF motifs that are targets of PP1 and Aurora B kinase and are most important for regulating the stability of microtubule attachments and maintaining metaphase I arrest. The region between amino acids 124 and 473 are required for two activities that are critical for accurate chromosome segregation in meiosis I, lateral microtubule attachments and bi-orientation of homologs.
Figures


References
-
- Bonner A.M., Hughes S.E., Chisholm J.A., Smith S.K., Slaughter B.D., Unruh J R., Collins K.A, Friederichs J.M., Florens L., Swanson S.K., Pelot M.C., Miller D.E., Washburn M.P., Jaspersen S.L., and Hawley R.S.. 2013. Binding of Drosophila Polo kinase to its regulator Matrimony is noncanonical and involves two separate functional domains. Proc Natl Acad Sci U S A. 110:E1222–1231. - PMC - PubMed
-
- Bonner A.M., Hughes S.E., and Hawley R.S.. 2020. Regulation of Polo Kinase by Matrimony Is Required for Cohesin Maintenance during Drosophila melanogaster Female Meiosis. Curr Biol. 30:715–722.e713. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials