

This is a preprint.
Mapping the Cellular Biogeography of Human Bone Marrow Niches Using Single-Cell Transcriptomics and Proteomic Imaging
- PMID: 38559168
- PMCID: PMC10979999
- DOI: 10.1101/2024.03.14.585083
Mapping the Cellular Biogeography of Human Bone Marrow Niches Using Single-Cell Transcriptomics and Proteomic Imaging
Update in
-
Mapping the cellular biogeography of human bone marrow niches using single-cell transcriptomics and proteomic imaging.Cell. 2024 Jun 6;187(12):3120-3140.e29. doi: 10.1016/j.cell.2024.04.013. Epub 2024 May 6. Cell. 2024. PMID: 38714197 Free PMC article.
Abstract
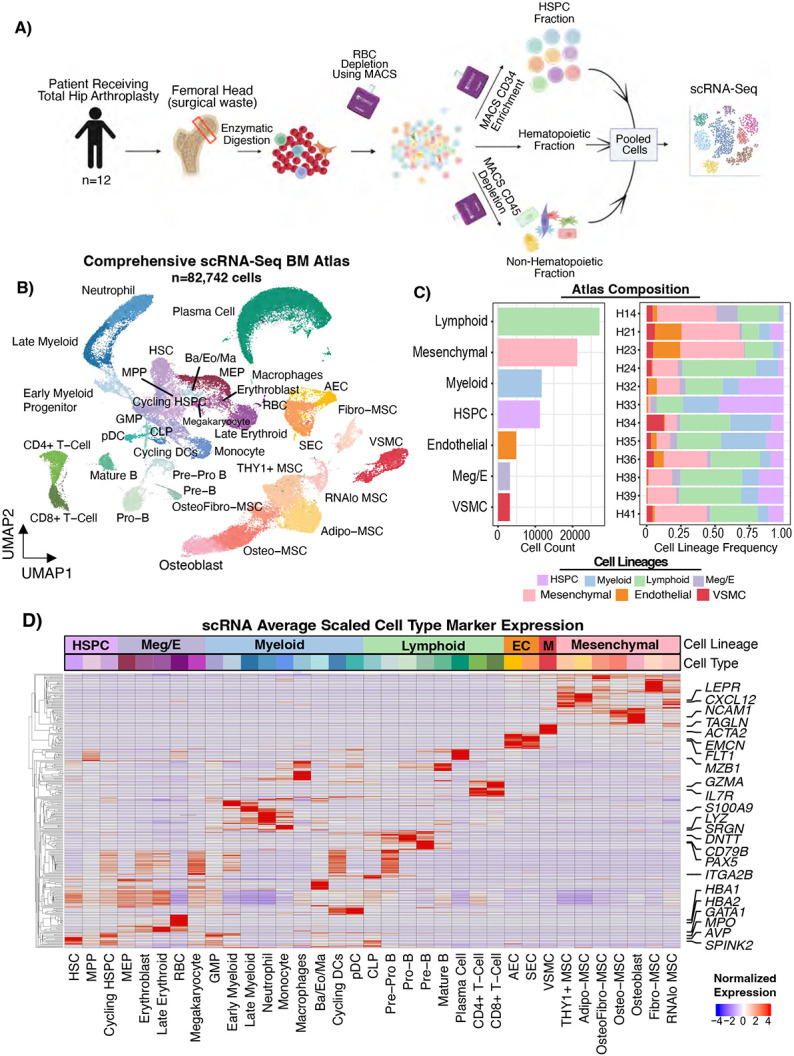
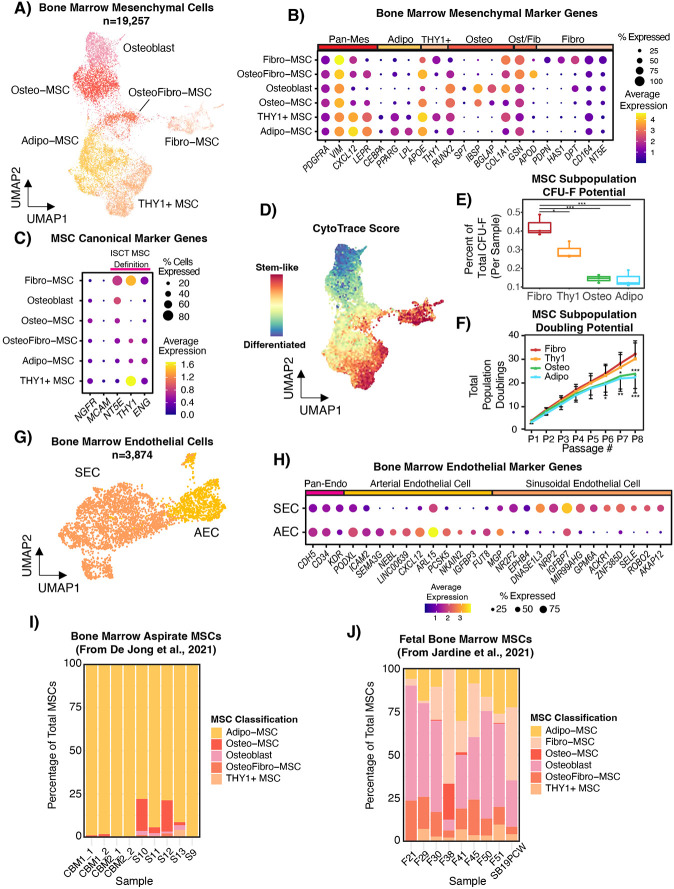
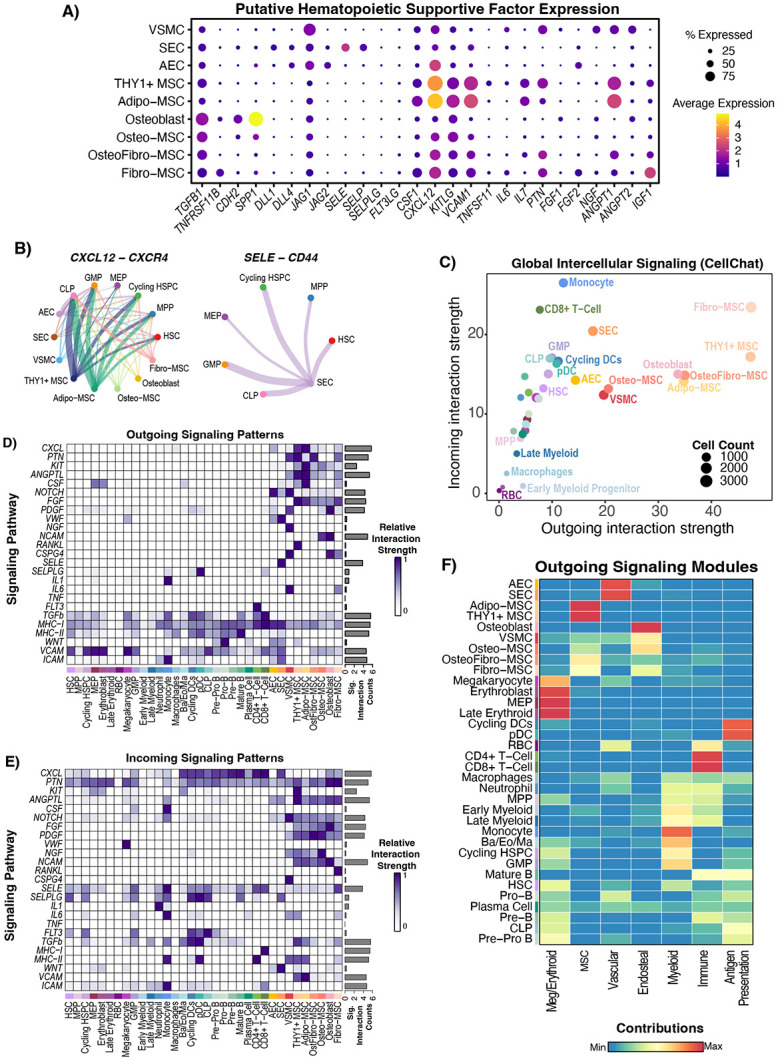
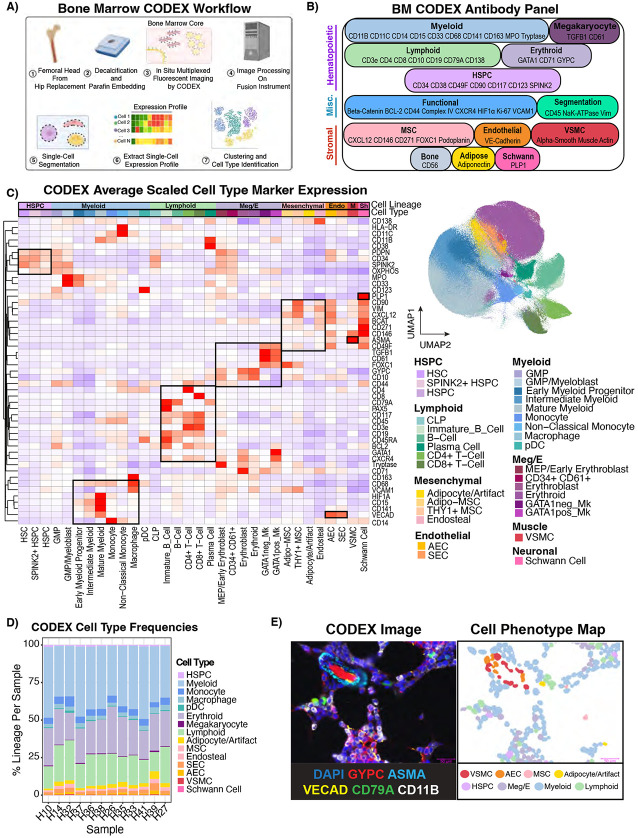
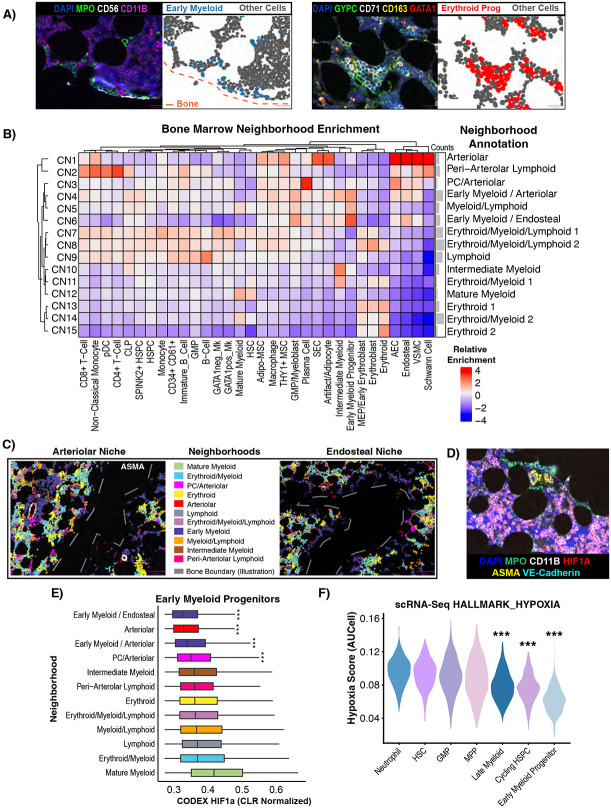
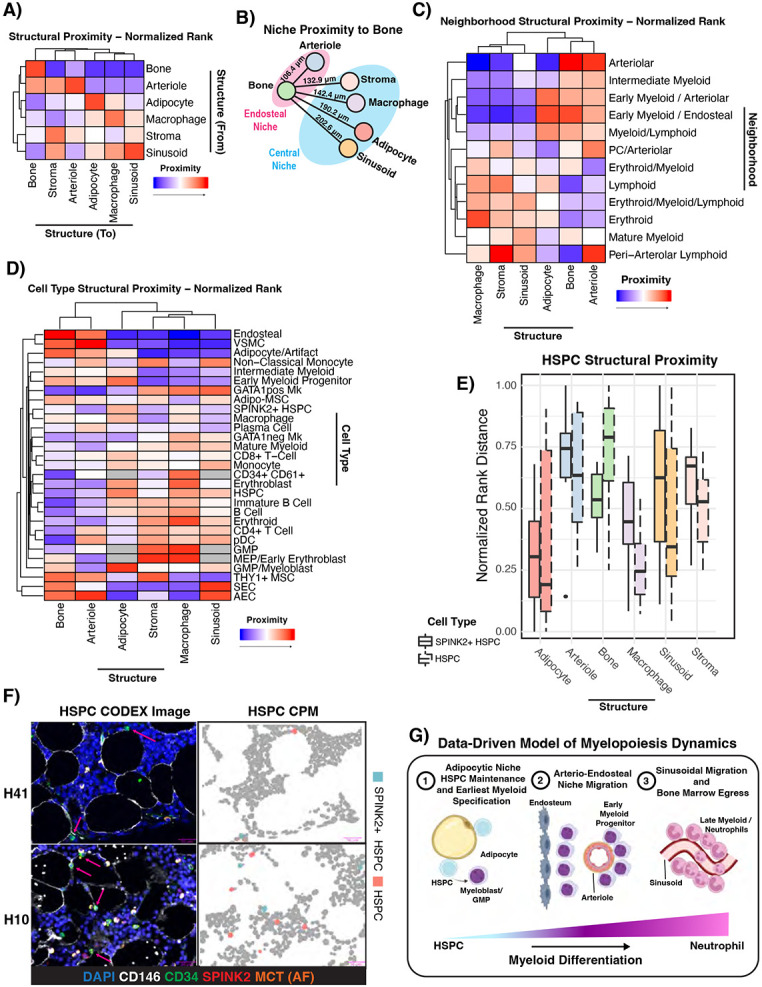
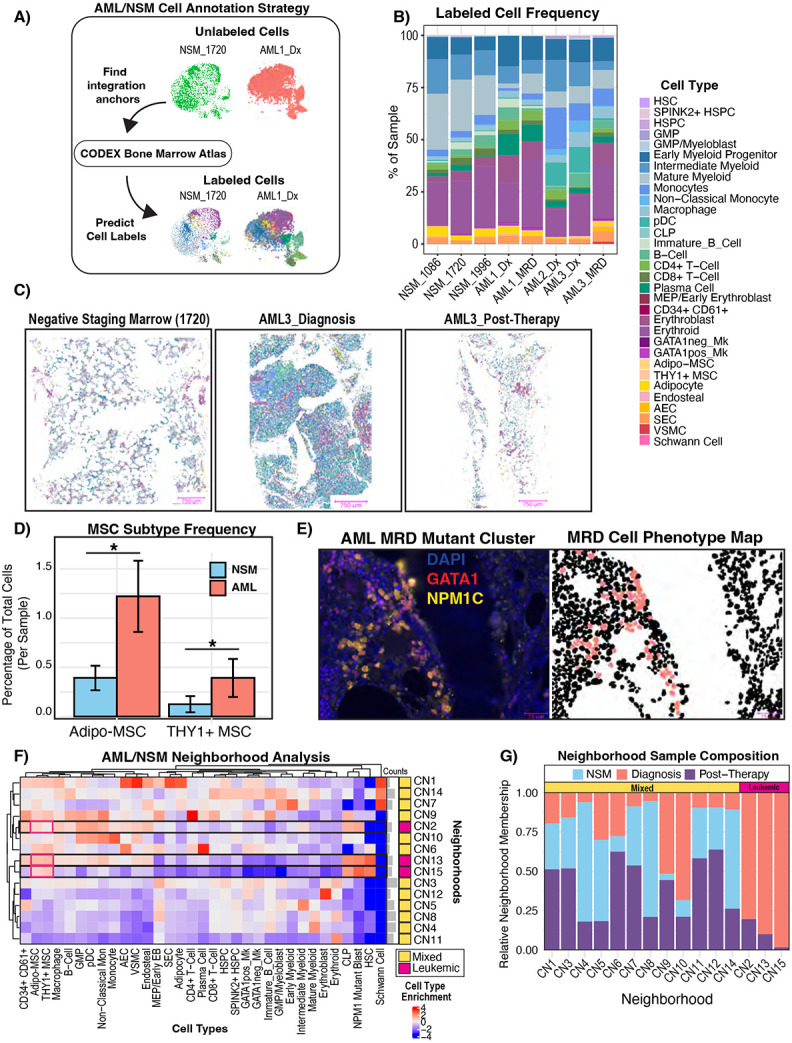
The bone marrow is the organ responsible for blood production. Diverse non-hematopoietic cells contribute essentially to hematopoiesis. However, these cells and their spatial organization remain largely uncharacterized as they have been technically challenging to study in humans. Here, we used fresh femoral head samples and performed single-cell RNA sequencing (scRNA-Seq) to profile 29,325 enriched non-hematopoietic bone marrow cells and discover nine transcriptionally distinct subtypes. We next employed CO-detection by inDEXing (CODEX) multiplexed imaging of 18 individuals, including both healthy and acute myeloid leukemia (AML) samples, to spatially profile over one million single cells with a novel 53-antibody panel. We discovered a relatively hyperoxygenated arterio-endosteal niche for early myelopoiesis, and an adipocytic, but not endosteal or perivascular, niche for early hematopoietic stem and progenitor cells. We used our atlas to predict cell type labels in new bone marrow images and used these predictions to uncover mesenchymal stromal cell (MSC) expansion and leukemic blast/MSC-enriched spatial neighborhoods in AML patient samples. Our work represents the first comprehensive, spatially-resolved multiomic atlas of human bone marrow and will serve as a reference for future investigation of cellular interactions that drive hematopoiesis.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Schofield R. (1978). The relationship between the spleen colony-forming cell and the haemopoietic stem cell. Blood Cells 4, 7–25. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials