

Sugar signals pedal the cell cycle!
- PMID: 38562561
- PMCID: PMC10982403
- DOI: 10.3389/fpls.2024.1354561
Sugar signals pedal the cell cycle!
Abstract
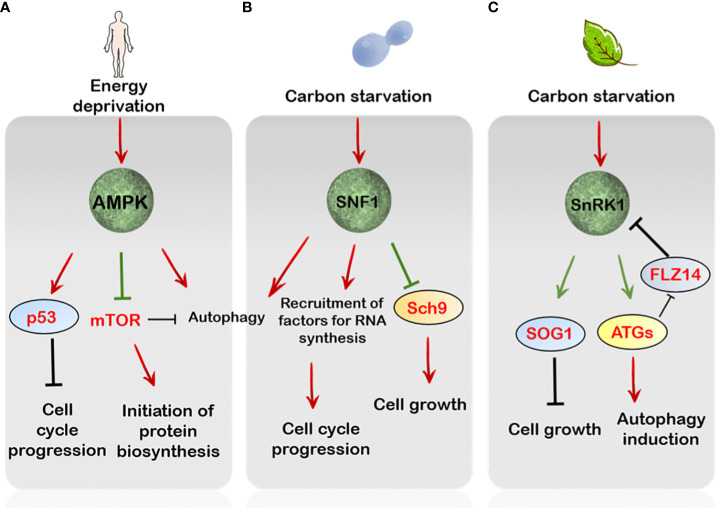
Cell cycle involves the sequential and reiterative progression of important events leading to cell division. Progression through a specific phase of the cell cycle is under the control of various factors. Since the cell cycle in multicellular eukaryotes responds to multiple extracellular mitogenic cues, its study in higher forms of life becomes all the more important. One such factor regulating cell cycle progression in plants is sugar signalling. Because the growth of organs depends on both cell growth and proliferation, sugars sensing and signalling are key control points linking sugar perception to regulation of downstream factors which facilitate these key developmental transitions. However, the basis of cell cycle control via sugars is intricate and demands exploration. This review deals with the information on sugar and TOR-SnRK1 signalling and how they manoeuvre various events of the cell cycle to ensure proper growth and development.
Keywords: CDK (cyclin-dependent kinase); SnRK1 kinase; glucose; sugar signalling; target of rapamycin (TOR).
Copyright © 2024 Rawat and Laxmi.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures


Similar articles
-
Cell cycle control by the target of rapamycin signalling pathway in plants.J Exp Bot. 2019 Apr 15;70(8):2275-2284. doi: 10.1093/jxb/erz140. J Exp Bot. 2019. PMID: 30918972 Review.
-
TOR and SnRK1 fine tune SPEECHLESS transcription and protein stability to optimize stomatal development in response to exogenously supplied sugar.New Phytol. 2022 Apr;234(1):107-121. doi: 10.1111/nph.17984. Epub 2022 Feb 8. New Phytol. 2022. PMID: 35060119
-
Interplay between cell growth and cell cycle in plants.J Exp Bot. 2014 Jun;65(10):2703-14. doi: 10.1093/jxb/ert354. Epub 2013 Nov 11. J Exp Bot. 2014. PMID: 24218325 Review.
-
Sugars and the speed of life-Metabolic signals that determine plant growth, development and death.Physiol Plant. 2022 Mar;174(2):e13656. doi: 10.1111/ppl.13656. Physiol Plant. 2022. PMID: 35243645 Free PMC article.
-
The critical roles of three sugar-related proteins (HXK, SnRK1, TOR) in regulating plant growth and stress responses.Hortic Res. 2024 Apr 4;11(6):uhae099. doi: 10.1093/hr/uhae099. eCollection 2024 Jun. Hortic Res. 2024. PMID: 38863993 Free PMC article.
Cited by
-
Transcriptome-based WGCNA reveals the molecular regulation of xylem plasticity in acclimation to drought and rewatering in mulberry.Front Plant Sci. 2024 Dec 18;15:1512645. doi: 10.3389/fpls.2024.1512645. eCollection 2024. Front Plant Sci. 2024. PMID: 39744604 Free PMC article.
-
C2H2 Zinc Finger Proteins GIS2 and ZFP8 Regulate Trichome Development via Hormone Signaling in Arabidopsis.Int J Mol Sci. 2025 Jul 27;26(15):7265. doi: 10.3390/ijms26157265. Int J Mol Sci. 2025. PMID: 40806397 Free PMC article.
-
The Cyclin-Dependent Kinase activity modulates the central carbon metabolism in maize during germination.Physiol Plant. 2025 Jan-Feb;177(1):e70119. doi: 10.1111/ppl.70119. Physiol Plant. 2025. PMID: 39956791 Free PMC article.
-
Modulating the activity of the APC/C regulator SISAMBA improves the sugar and antioxidant content of tomato fruits.Plant Biotechnol J. 2025 Sep;23(9):3540-3560. doi: 10.1111/pbi.70149. Epub 2025 Jun 8. Plant Biotechnol J. 2025. PMID: 40483582 Free PMC article.
References
Publication types
LinkOut - more resources
Full Text Sources