

Gradients of bacteria in the oceanic water column reveal finely-resolved vertical distributions
- PMID: 38564528
- PMCID: PMC10986988
- DOI: 10.1371/journal.pone.0298139
Gradients of bacteria in the oceanic water column reveal finely-resolved vertical distributions
Abstract
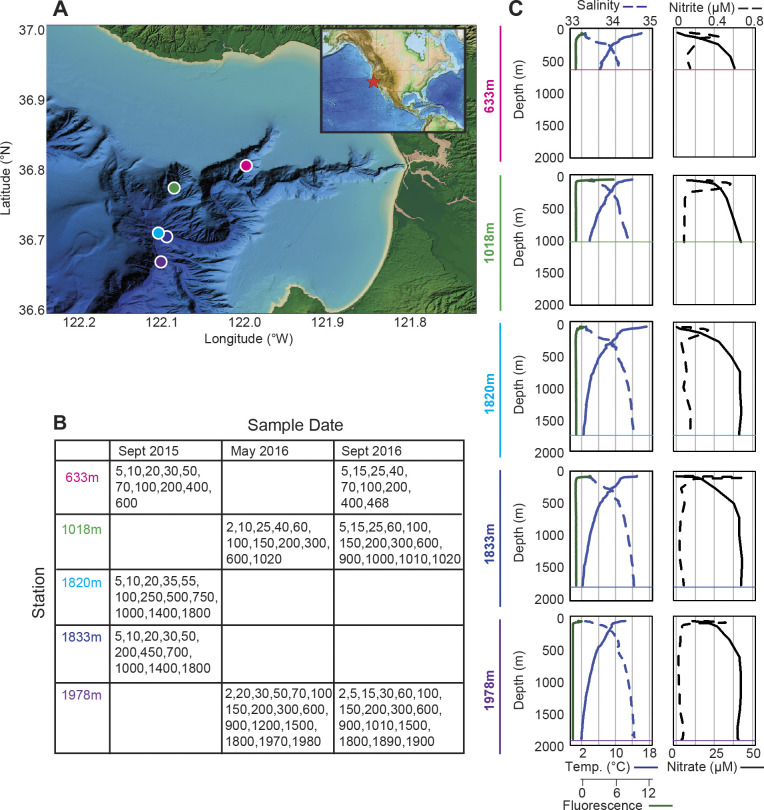
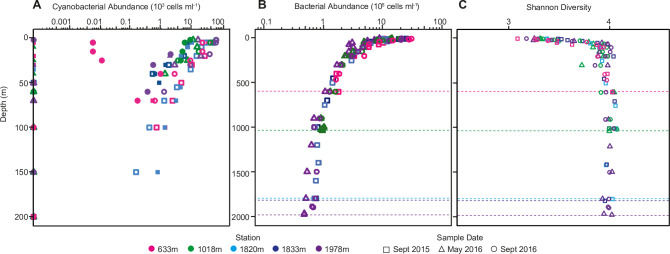
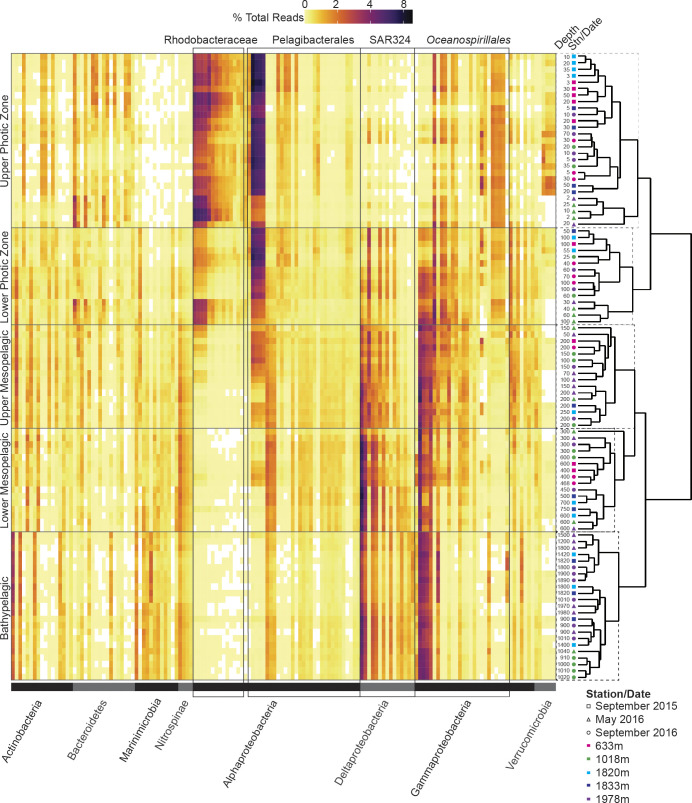
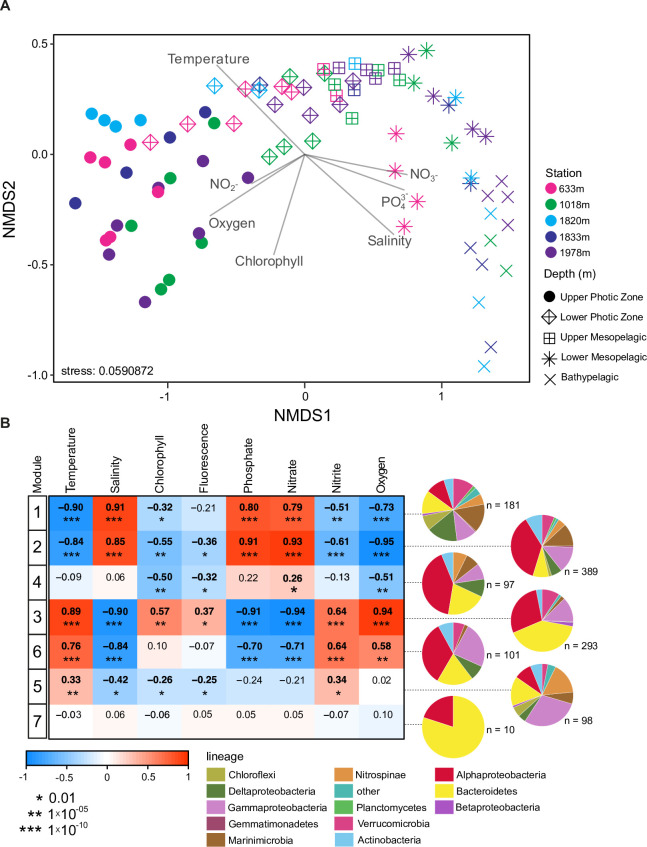
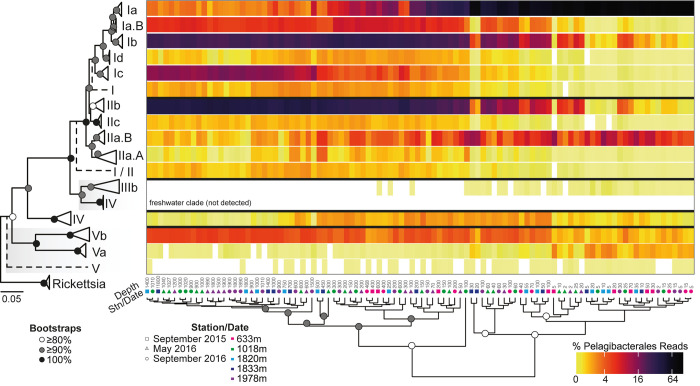
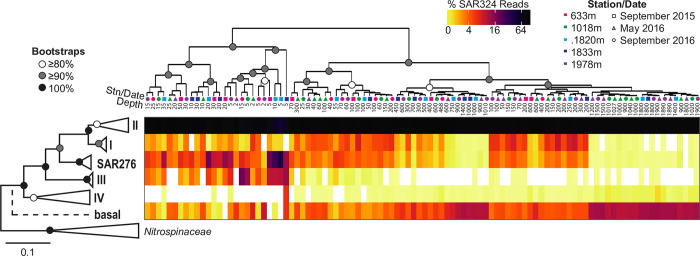
Bacterial communities directly influence ecological processes in the ocean, and depth has a major influence due to the changeover in primary energy sources between the sunlit photic zone and dark ocean. Here, we examine the abundance and diversity of bacteria in Monterey Bay depth profiles collected from the surface to just above the sediments (e.g., 2000 m). Bacterial abundance in these Pacific Ocean samples decreased by >1 order of magnitude, from 1.22 ±0.69 ×106 cells ml-1 in the variable photic zone to 1.44 ± 0.25 ×105 and 6.71 ± 1.23 ×104 cells ml-1 in the mesopelagic and bathypelagic, respectively. V1-V2 16S rRNA gene profiling showed diversity increased sharply between the photic and mesopelagic zones. Weighted Gene Correlation Network Analysis clustered co-occurring bacterial amplicon sequence variants (ASVs) into seven subnetwork modules, of which five strongly correlated with depth-related factors. Within surface-associated modules there was a clear distinction between a 'copiotrophic' module, correlating with chlorophyll and dominated by e.g., Flavobacteriales and Rhodobacteraceae, and an 'oligotrophic' module dominated by diverse Oceanospirillales (such as uncultured JL-ETNP-Y6, SAR86) and Pelagibacterales. Phylogenetic reconstructions of Pelagibacterales and SAR324 using full-length 16S rRNA gene data revealed several additional subclades, expanding known microdiversity within these abundant lineages, including new Pelagibacterales subclades Ia.B, Id, and IIc, which comprised 4-10% of amplicons depending on the subclade and depth zone. SAR324 and Oceanospirillales dominated in the mesopelagic, with SAR324 clade II exhibiting its highest relative abundances (17±4%) in the lower mesopelagic (300-750 m). The two newly-identified SAR324 clades showed highest relative abundances in the photic zone (clade III), while clade IV was extremely low in relative abundance, but present across dark ocean depths. Hierarchical clustering placed microbial communities from 900 m samples with those from the bathypelagic, where Marinimicrobia was distinctively relatively abundant. The patterns resolved herein, through high resolution and statistical replication, establish baselines for marine bacterial abundance and taxonomic distributions across the Monterey Bay water column, against which future change can be assessed.
Copyright: © 2024 Harbeitner et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The author have declared that no competing interests exist.
Figures
References
-
- Sunagawa S, Coelho LP, Chaffron S, Kultima JR, Labadie K, Salazar G, et al. Structure and function of the global ocean microbiome. Science (1979) 2015; 348: 1261359. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
