

EP300-ZNF384 transactivates IL3RA to promote the progression of B-cell acute lymphoblastic leukemia
- PMID: 38566191
- PMCID: PMC10986138
- DOI: 10.1186/s12964-024-01596-9
EP300-ZNF384 transactivates IL3RA to promote the progression of B-cell acute lymphoblastic leukemia
Abstract
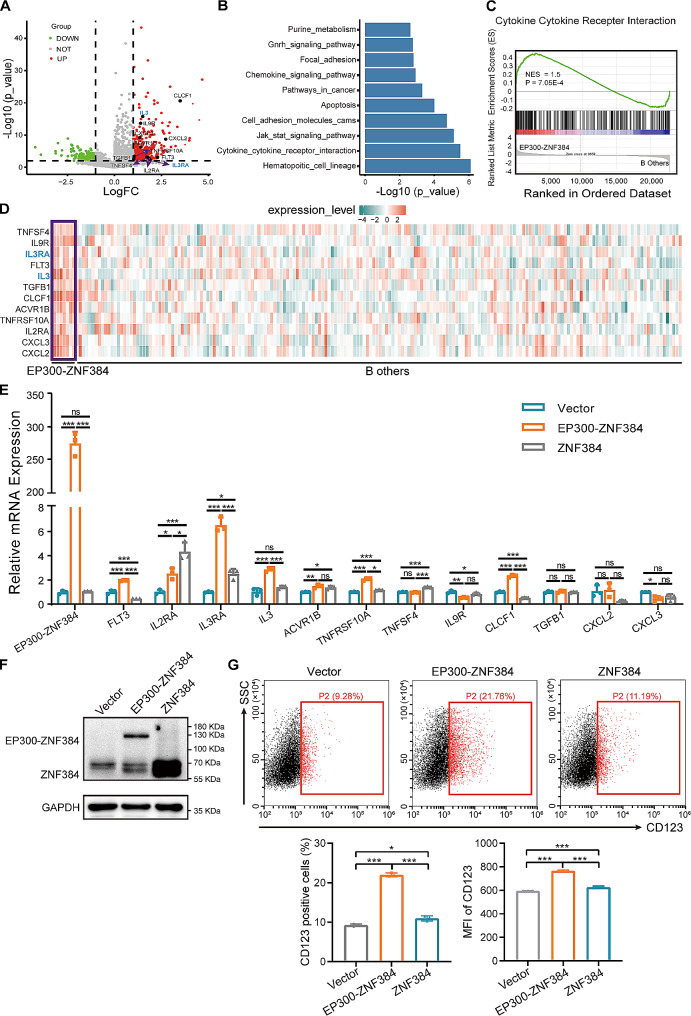
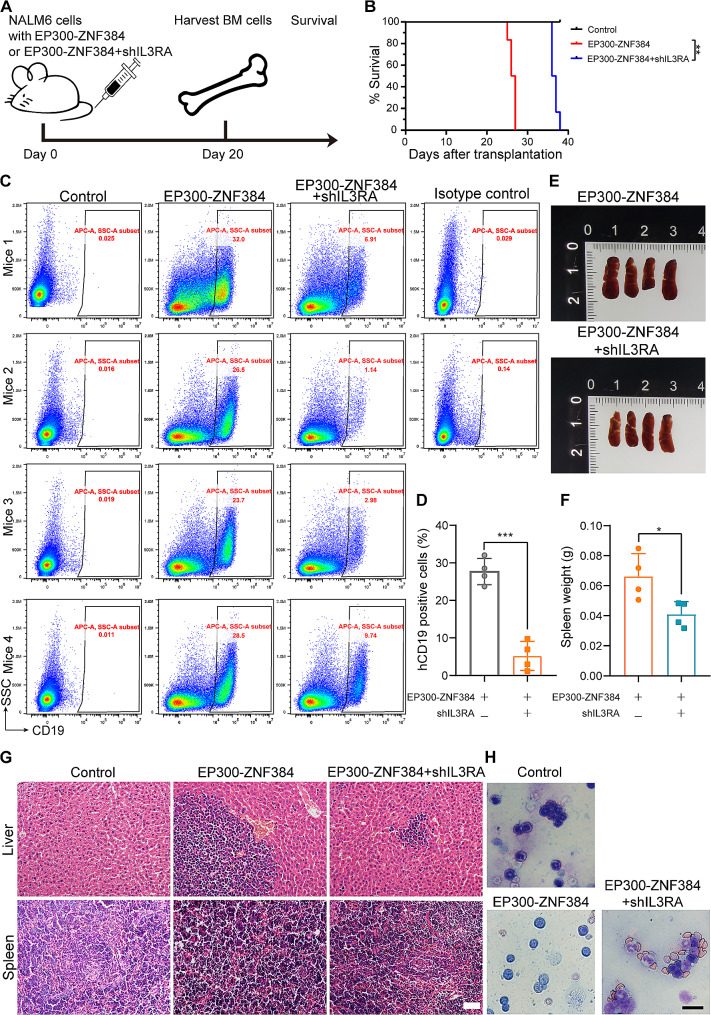
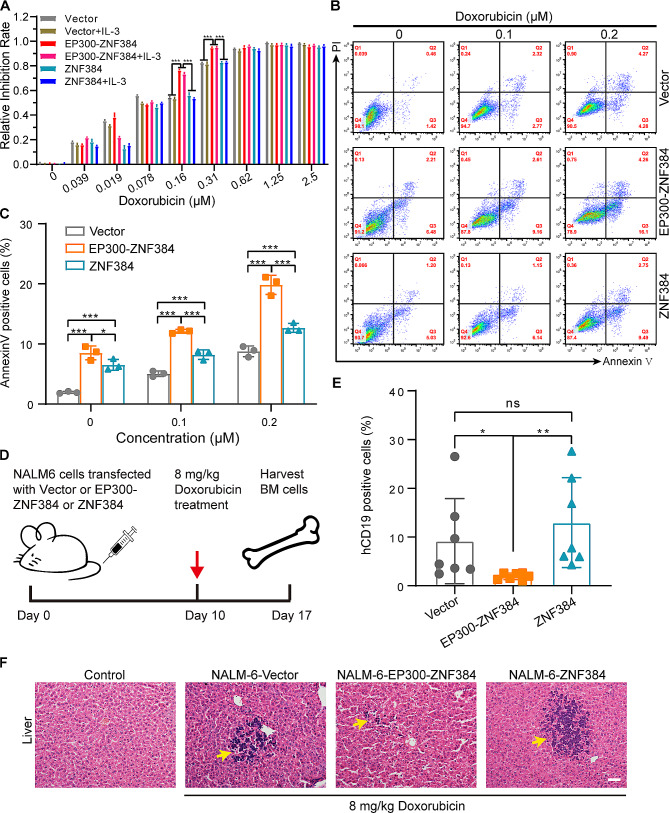
The EP300-ZNF384 fusion gene is an oncogenic driver in B-cell acute lymphoblastic leukemia (B-ALL). In the present study, we demonstrated that EP300-ZNF384 substantially induces the transcription of IL3RA and the expression of IL3Rα (CD123) on B-ALL cell membranes. Interleukin 3 (IL-3) supplementation promotes the proliferation of EP300-ZNF348-positive B-ALL cells by activating STAT5. Conditional knockdown of IL3RA in EP300-ZF384-positive cells inhibited the proliferation in vitro, and induced a significant increase in overall survival of mice, which is attributed to impaired propagation ability of leukemia cells. Mechanistically, the EP300-ZNF384 fusion protein transactivates the promoter activity of IL3RA by binding to an A-rich sequence localized at -222/-234 of IL3RA. Furthermore, forced EP300-ZNF384 expression induces the expression of IL3Rα on cell membranes and the secretion of IL-3 in CD19-positive B precursor cells derived from healthy individuals. Doxorubicin displayed a selective killing of EP300-ZNF384-positive B-ALL cells in vitro and in vivo. Collectively, we identify IL3RA as a direct downstream target of EP300-ZNF384, suggesting CD123 is a potent biomarker for EP300-ZNF384-driven B-ALL. Targeting CD123 may be a novel therapeutic approach to EP300-ZNF384-positive patients, alternative or, more likely, complementary to standard chemotherapy regimen in clinical setting.
Keywords: B-cell acute lymphoblastic leukemia; EP300-ZNF384; IL-3; IL3RA; Leukemogenesis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
EP300-ZNF384 fusion gene product up-regulates GATA3 gene expression and induces hematopoietic stem cell gene expression signature in B-cell precursor acute lymphoblastic leukemia cells.Int J Hematol. 2017 Aug;106(2):269-281. doi: 10.1007/s12185-017-2220-6. Epub 2017 Apr 4. Int J Hematol. 2017. PMID: 28378055
-
Detection of EP300-ZNF384 fusion in patients with acute lymphoblastic leukemia using RNA fusion gene panel sequencing.Ann Hematol. 2020 Nov;99(11):2611-2617. doi: 10.1007/s00277-020-04251-8. Epub 2020 Sep 26. Ann Hematol. 2020. PMID: 32980888 Free PMC article. Clinical Trial.
-
IKAROS and CK2 regulate expression of BCL-XL and chemosensitivity in high-risk B-cell acute lymphoblastic leukemia.Blood. 2020 Sep 24;136(13):1520-1534. doi: 10.1182/blood.2019002655. Blood. 2020. PMID: 32396934 Free PMC article.
-
PEG-asparaginase treatment regimens for acute lymphoblastic leukaemia in children: a network meta-analysis.Cochrane Database Syst Rev. 2023 May 31;5(5):CD014570. doi: 10.1002/14651858.CD014570.pub2. Cochrane Database Syst Rev. 2023. PMID: 37260073 Free PMC article.
-
Taxane monotherapy regimens for the treatment of recurrent epithelial ovarian cancer.Cochrane Database Syst Rev. 2022 Jul 12;7(7):CD008766. doi: 10.1002/14651858.CD008766.pub3. Cochrane Database Syst Rev. 2022. PMID: 35866378 Free PMC article.
Cited by
-
Transcriptomic profiling in canine B-cell lymphoma supports a synergistic effect of BTK and PI3K inhibitors.Front Vet Sci. 2025 Apr 25;12:1577028. doi: 10.3389/fvets.2025.1577028. eCollection 2025. Front Vet Sci. 2025. PMID: 40351764 Free PMC article.
-
The role of chemokines and interleukins in acute lymphoblastic leukemia: a systematic review.J Appl Biomed. 2024 Dec;22(4):165-184. doi: 10.32725/jab.2024.024. Epub 2024 Dec 4. J Appl Biomed. 2024. PMID: 40033805
References
-
- Hirabayashi S, Ohki K, Nakabayashi K, Ichikawa H, Momozawa Y, Okamura K, et al. ZNF384-related fusion genes define a subgroup of childhood B-cell precursor acute lymphoblastic leukemia with a characteristic immunotype. Haematologica. 2017;102(1):118–29. doi: 10.3324/haematol.2016.151035. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 508010/the Youth Talent Cultivation Fund Project of Dalian Medical University
- JYTZD2023041/the Basic Research project of the Educational Department of Liaoning Province
- 2023JH6/100100019/the Central Guidance on Local Science and Technology Development Fund of Liaoning Province
- XLYC1902036/the Science and Technology Innovation Leading Talent Program of Liaoning Province
- 2019J12SN56/the Basic Research on the Application of Dalian Innovation Fund
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
