

Structure-Toxicity Relationship in Intermediate Fibrils from α-Synuclein Condensates
- PMID: 38567991
- PMCID: PMC11027145
- DOI: 10.1021/jacs.3c14703
Structure-Toxicity Relationship in Intermediate Fibrils from α-Synuclein Condensates
Abstract
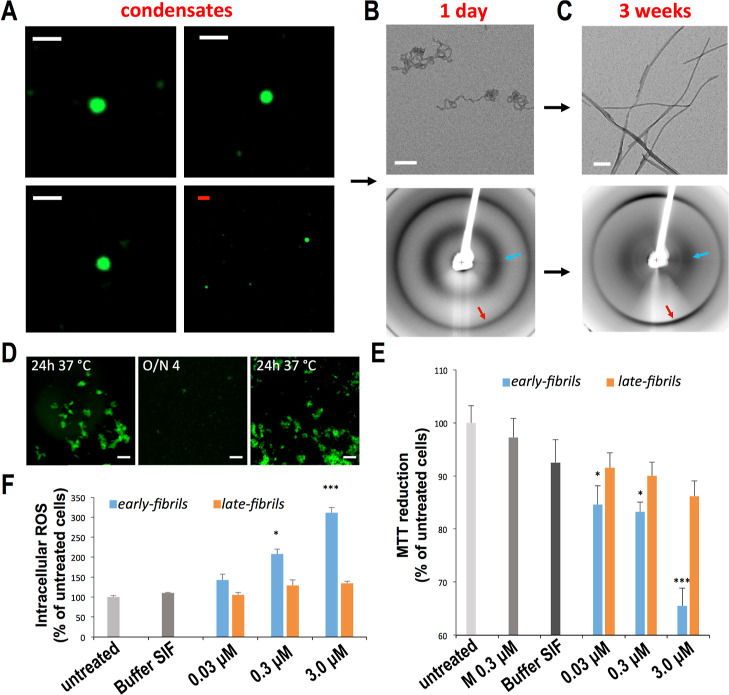
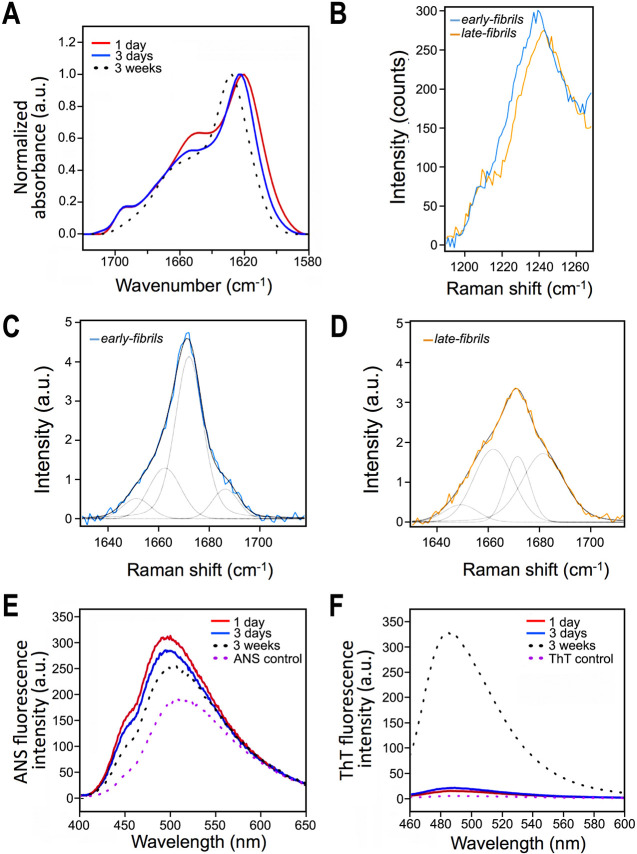
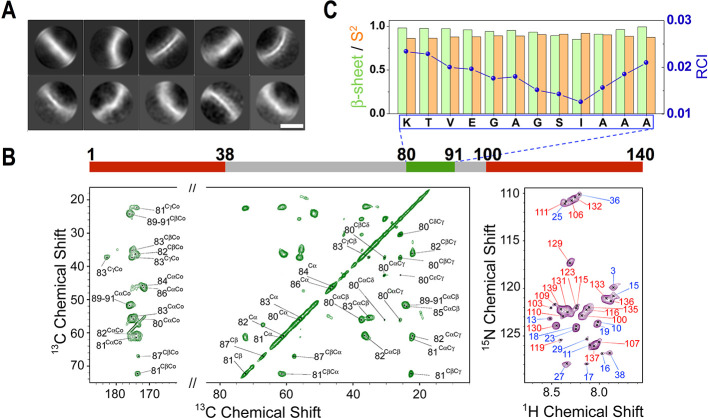
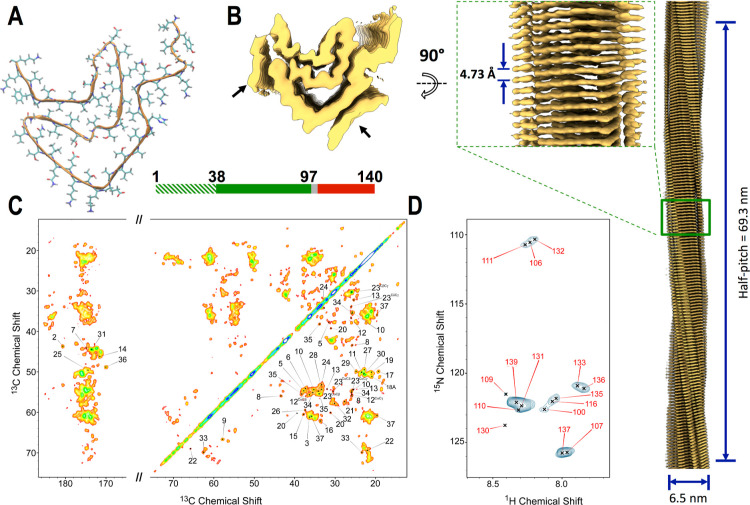
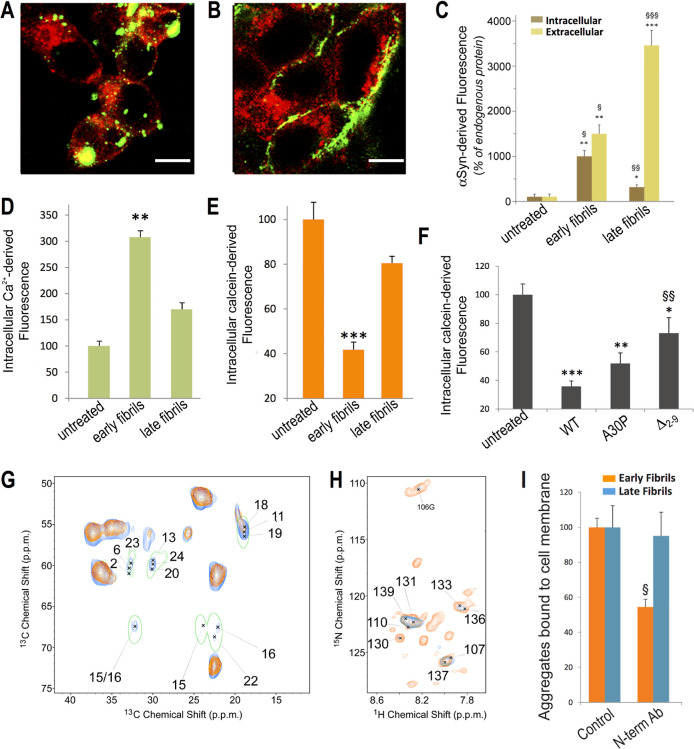
The aberrant aggregation of α-synuclein (αS) into amyloid fibrils is associated with a range of highly debilitating neurodegenerative conditions, including Parkinson's disease. Although the structural properties of mature amyloids of αS are currently understood, the nature of transient protofilaments and fibrils that appear during αS aggregation remains elusive. Using solid-state nuclear magnetic resonance (ssNMR), cryogenic electron microscopy (cryo-EM), and biophysical methods, we here characterized intermediate amyloid fibrils of αS forming during the aggregation from liquid-like spherical condensates to mature amyloids adopting the structure of pathologically observed aggregates. These transient amyloid intermediates, which induce significant levels of cytotoxicity when incubated with neuronal cells, were found to be stabilized by a small core in an antiparallel β-sheet conformation, with a disordered N-terminal region of the protein remaining available to mediate membrane binding. In contrast, mature amyloids that subsequently appear during the aggregation showed different structural and biological properties, including low levels of cytotoxicity, a rearranged structured core embedding also the N-terminal region, and a reduced propensity to interact with the membrane. The characterization of these two fibrillar forms of αS, and the use of antibodies and designed mutants, enabled us to clarify the role of critical structural elements endowing intermediate amyloid species with the ability to interact with membranes and induce cytotoxicity.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Fusco G.; Chen S. W.; Williamson P. T. F.; Cascella R.; Perni M.; Jarvis J. A.; Cecchi C.; Vendruscolo M.; Chiti F.; Cremades N.; Ying L.; Dobson C. M.; De Simone A. Structural basis of membrane disruption and cellular toxicity by α-synuclein oligomers. Science 2017, 358 (6369), 1440–1443. 10.1126/science.aan6160. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
