

The variation and evolution of complete human centromeres
- PMID: 38570684
- PMCID: PMC11062924
- DOI: 10.1038/s41586-024-07278-3
The variation and evolution of complete human centromeres
Abstract
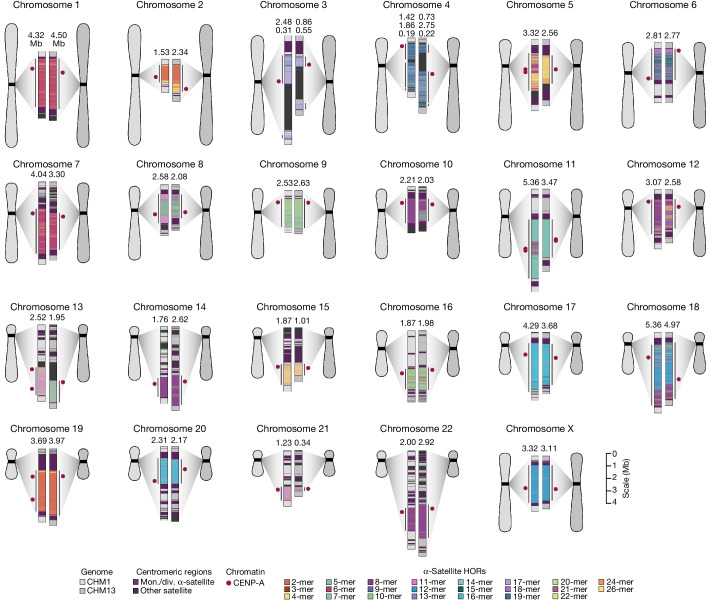
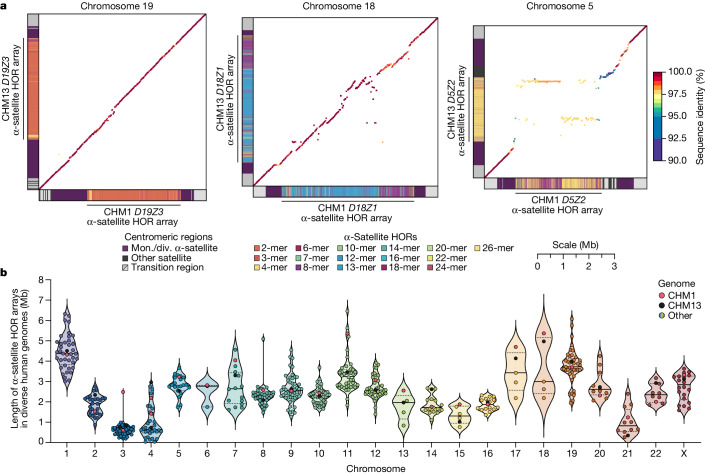
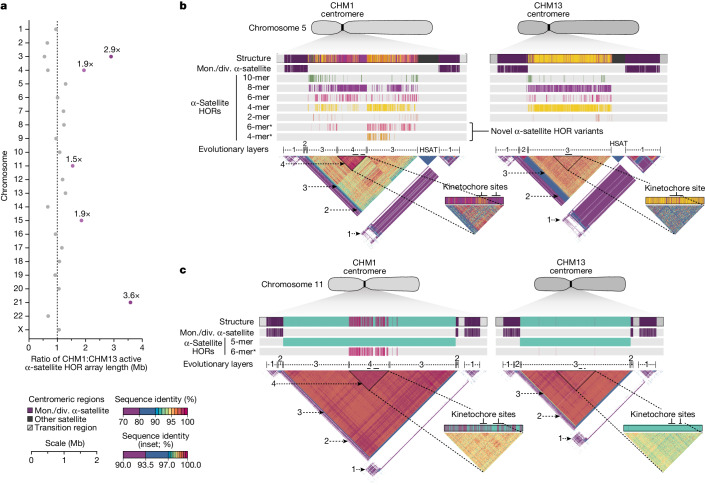
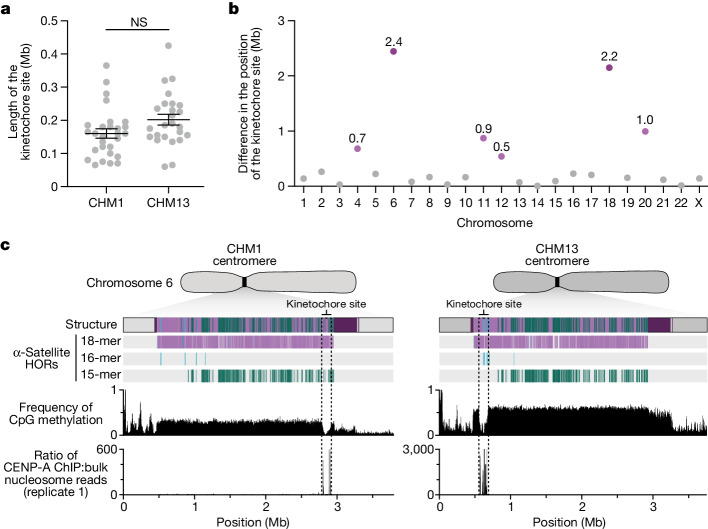
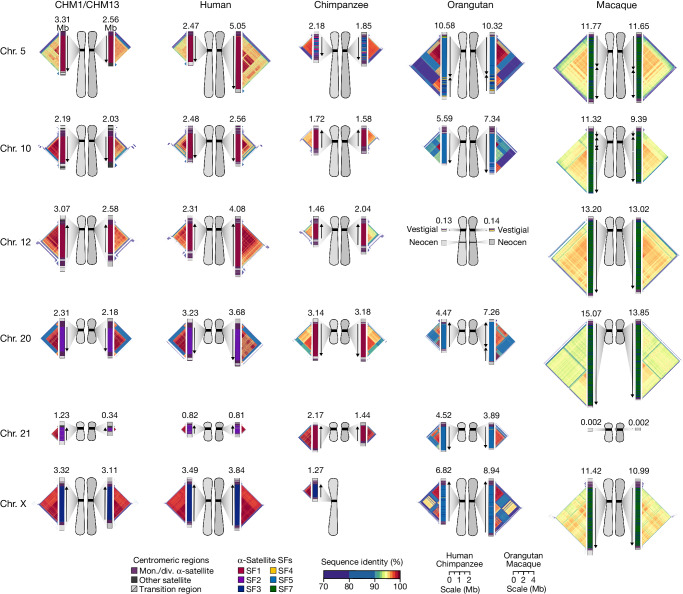
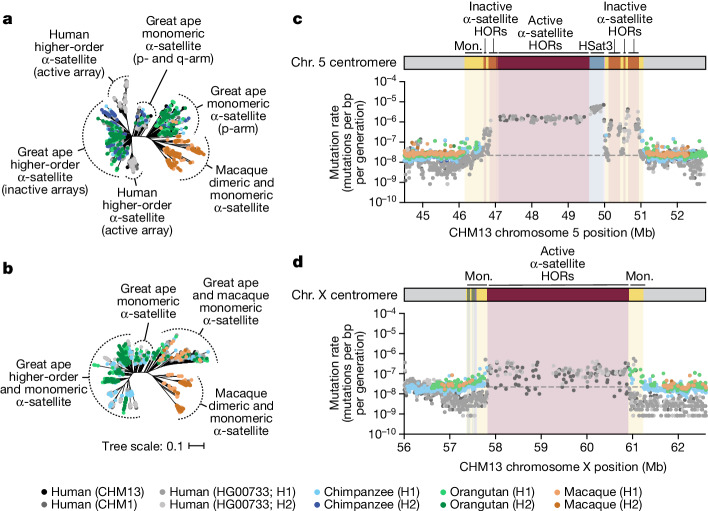
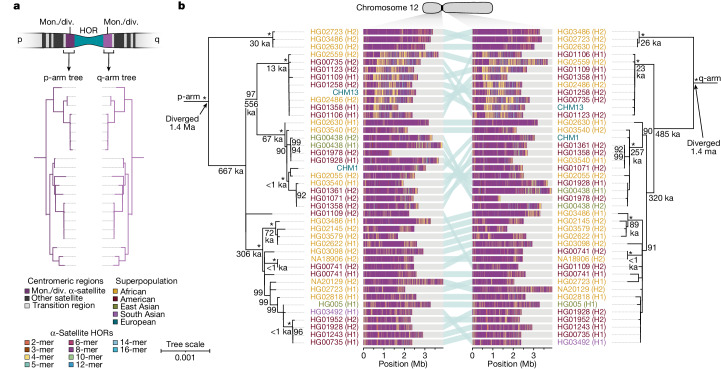
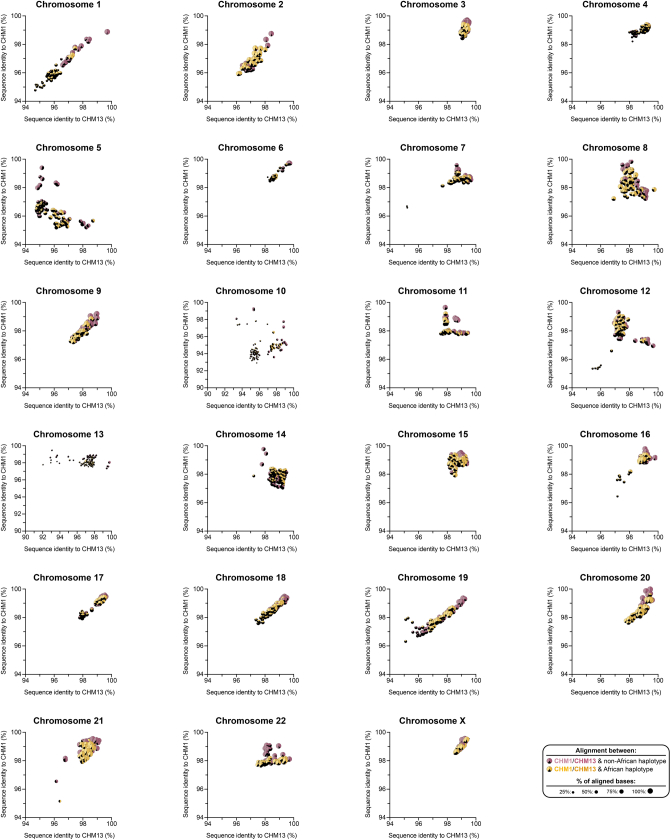
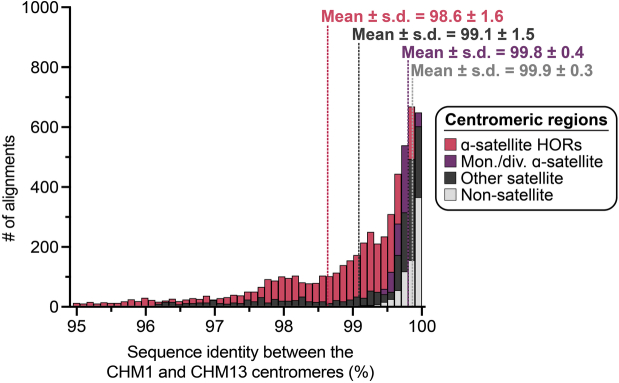
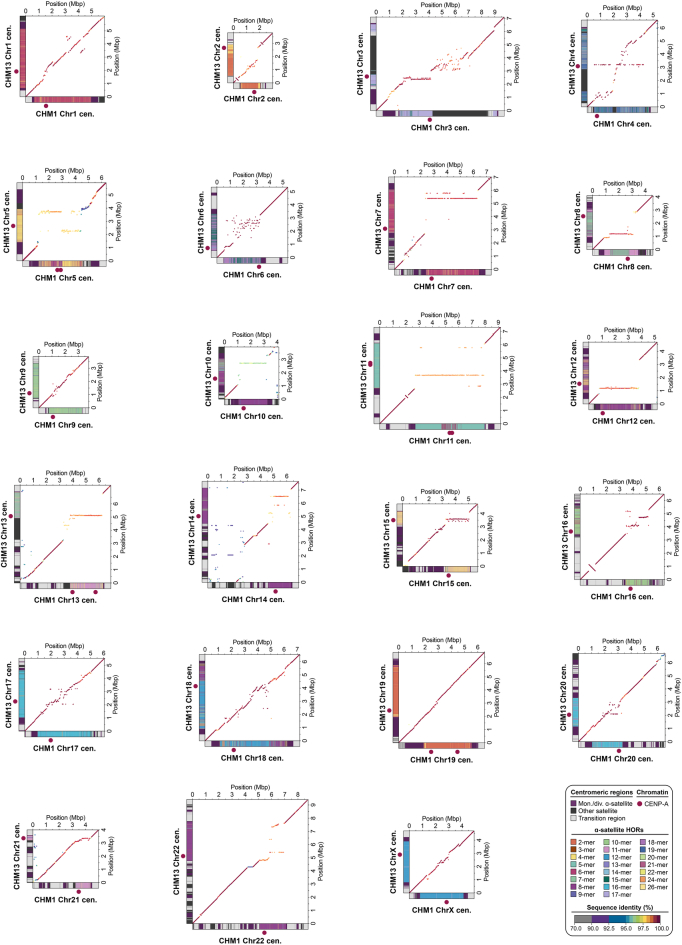
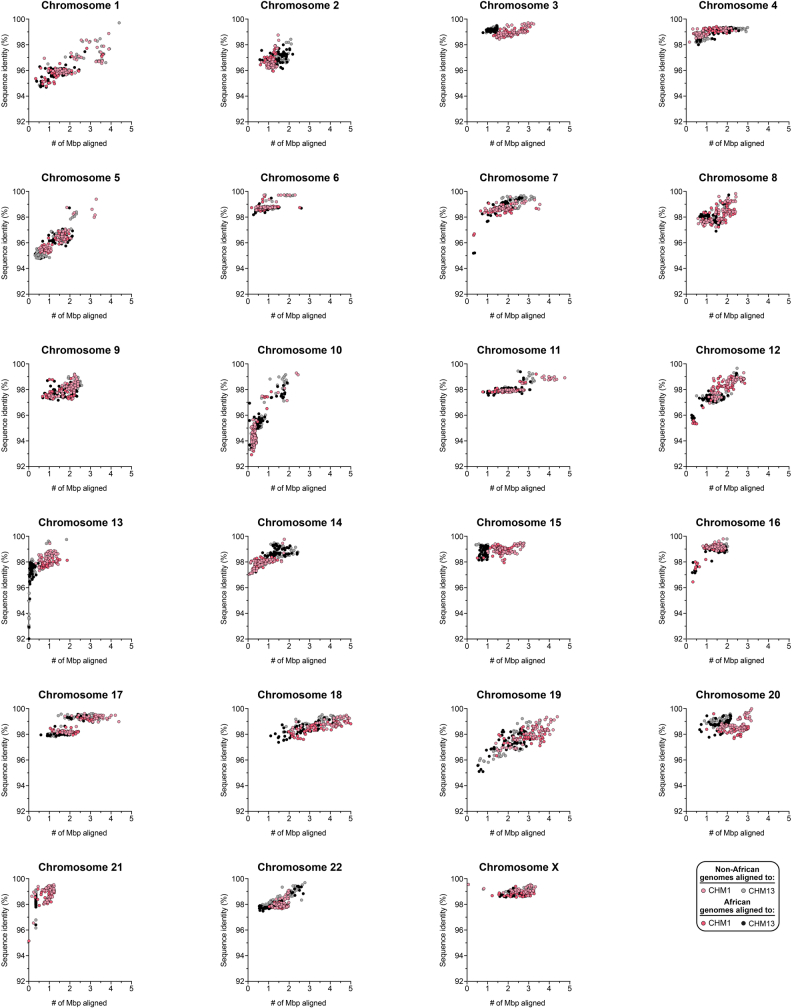
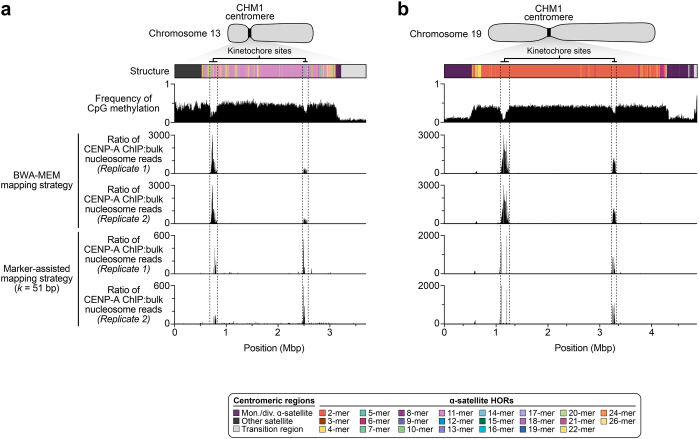
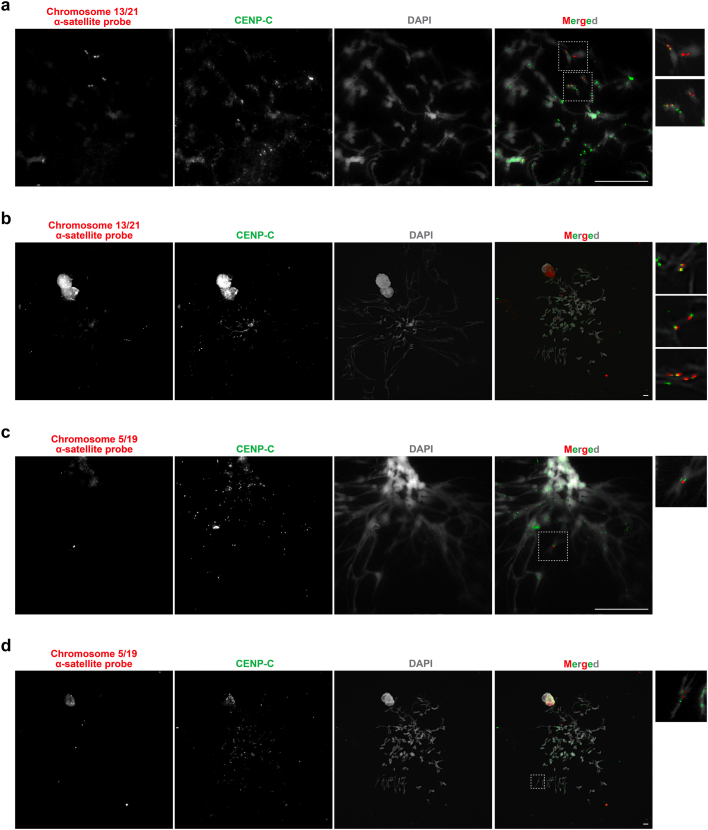
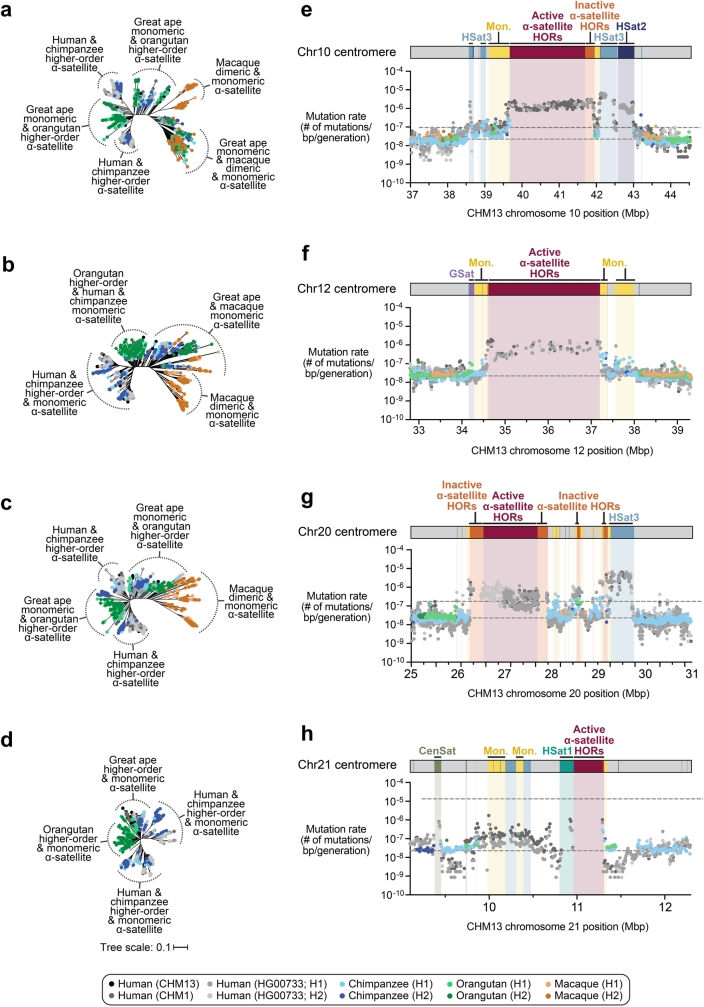
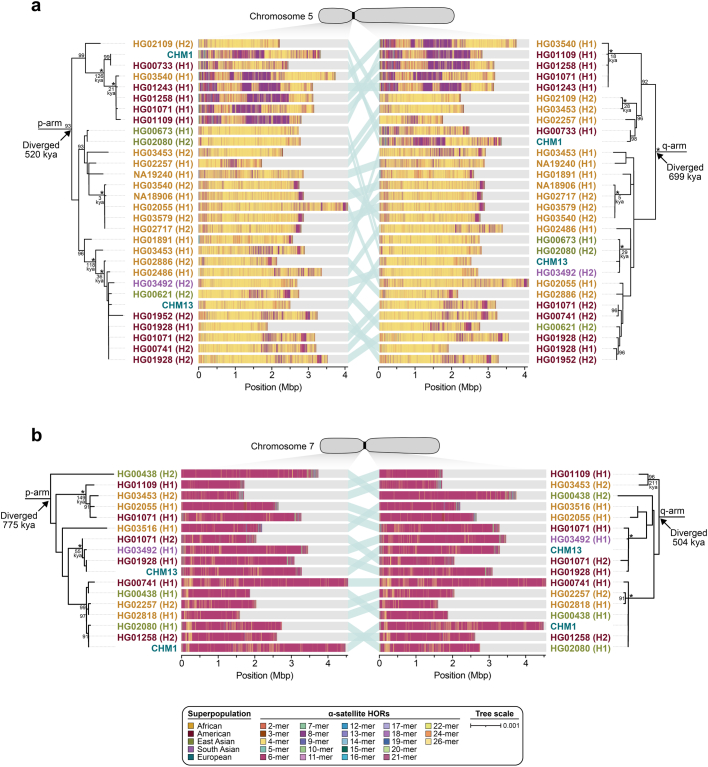
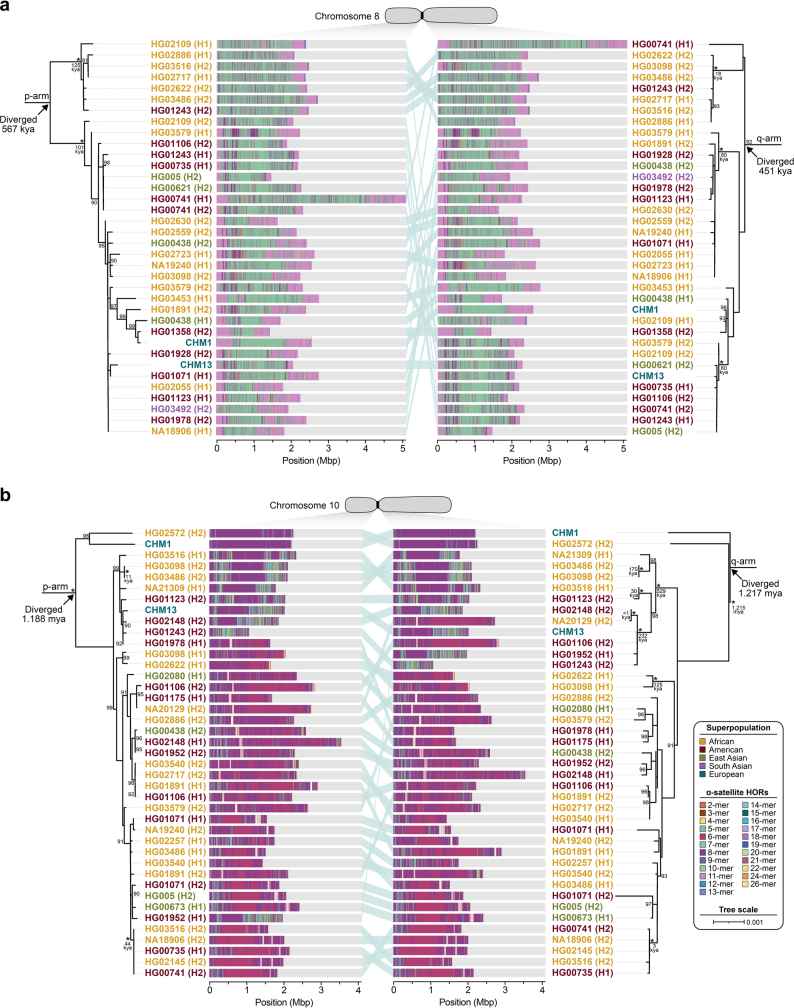
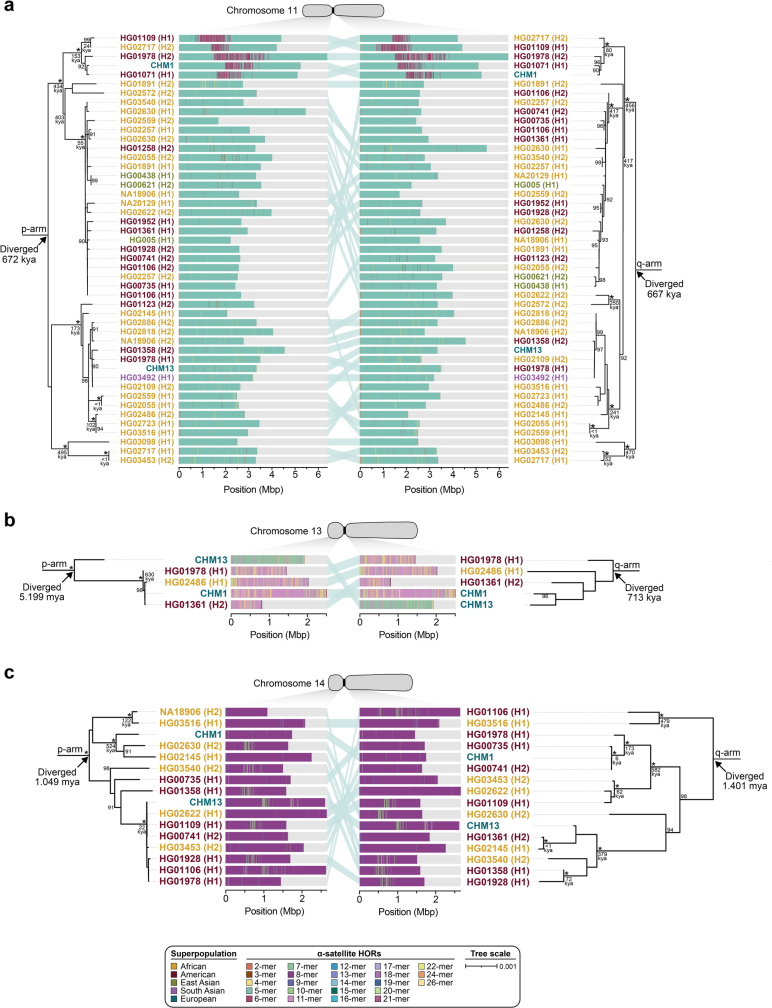
Human centromeres have been traditionally very difficult to sequence and assemble owing to their repetitive nature and large size1. As a result, patterns of human centromeric variation and models for their evolution and function remain incomplete, despite centromeres being among the most rapidly mutating regions2,3. Here, using long-read sequencing, we completely sequenced and assembled all centromeres from a second human genome and compared it to the finished reference genome4,5. We find that the two sets of centromeres show at least a 4.1-fold increase in single-nucleotide variation when compared with their unique flanks and vary up to 3-fold in size. Moreover, we find that 45.8% of centromeric sequence cannot be reliably aligned using standard methods owing to the emergence of new α-satellite higher-order repeats (HORs). DNA methylation and CENP-A chromatin immunoprecipitation experiments show that 26% of the centromeres differ in their kinetochore position by >500 kb. To understand evolutionary change, we selected six chromosomes and sequenced and assembled 31 orthologous centromeres from the common chimpanzee, orangutan and macaque genomes. Comparative analyses reveal a nearly complete turnover of α-satellite HORs, with characteristic idiosyncratic changes in α-satellite HORs for each species. Phylogenetic reconstruction of human haplotypes supports limited to no recombination between the short (p) and long (q) arms across centromeres and reveals that novel α-satellite HORs share a monophyletic origin, providing a strategy to estimate the rate of saltatory amplification and mutation of human centromeric DNA.
© 2024. The Author(s).
Conflict of interest statement
S.N. is an employee of Oxford Nanopore Technologies. S.K. has received travel funds to speak at events hosted by Oxford Nanopore Technologies. E.E.E. is a scientific advisory board member of Variant Bio. The other authors declare no competing interests.
Figures

Update of
-
The variation and evolution of complete human centromeres.bioRxiv [Preprint]. 2023 May 30:2023.05.30.542849. doi: 10.1101/2023.05.30.542849. bioRxiv. 2023. Update in: Nature. 2024 May;629(8010):136-145. doi: 10.1038/s41586-024-07278-3. PMID: 37398417 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
