

Neutrophil-mediated innate immune resistance to bacterial pneumonia is dependent on Tet2 function
- PMID: 38573824
- PMCID: PMC11142737
- DOI: 10.1172/JCI171002
Neutrophil-mediated innate immune resistance to bacterial pneumonia is dependent on Tet2 function
Abstract
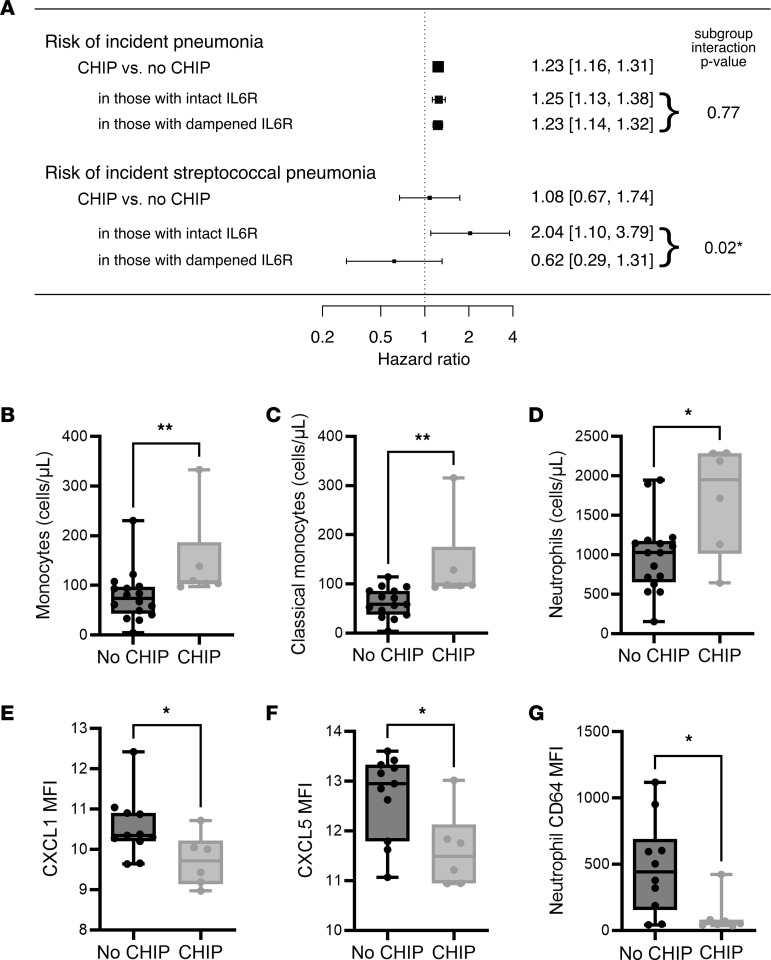
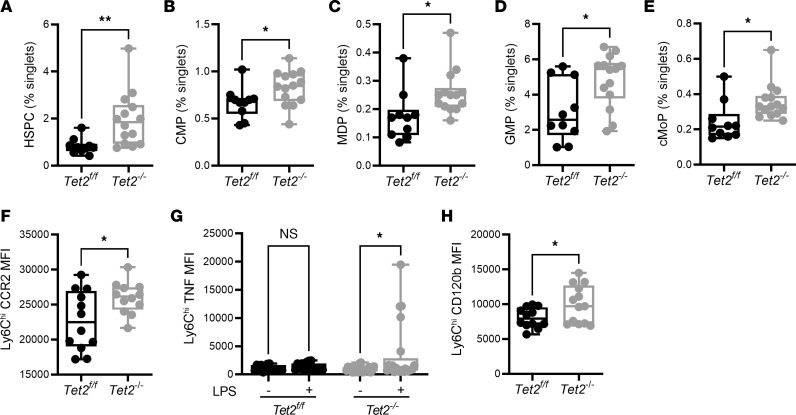
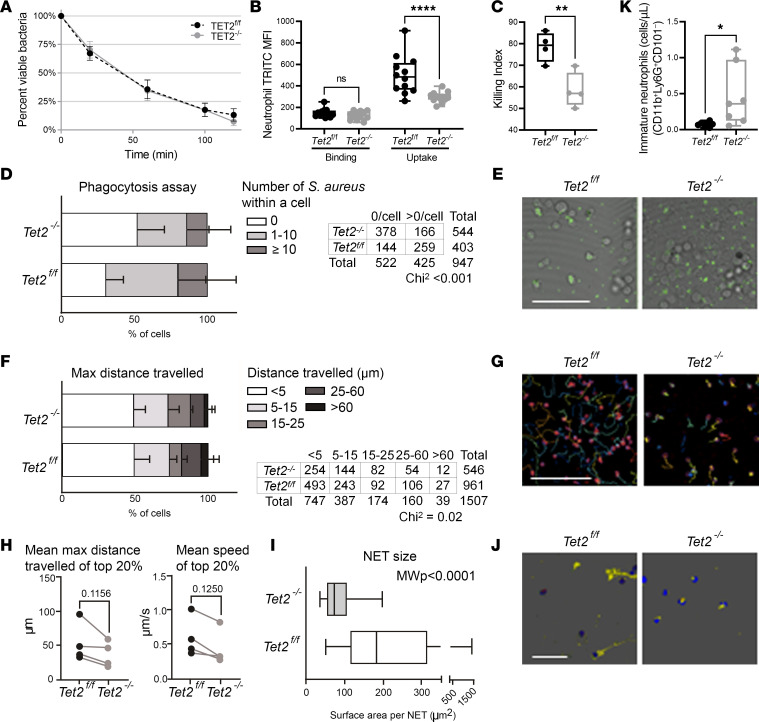
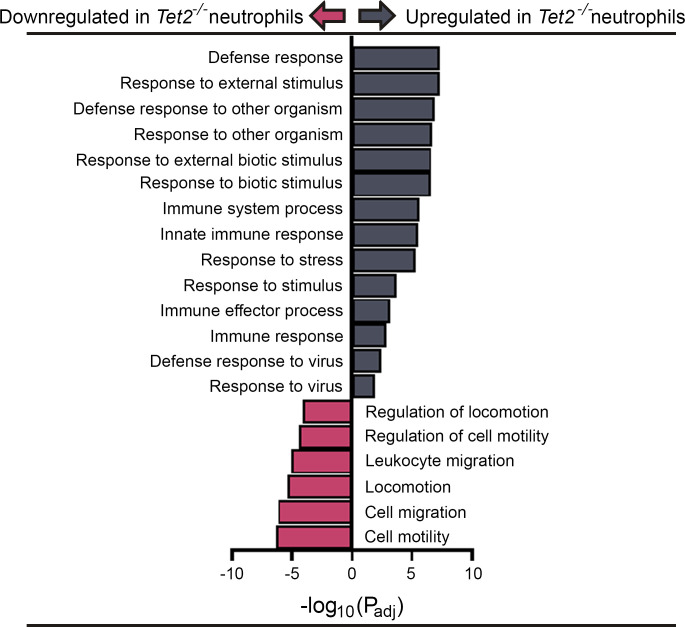
Individuals with clonal hematopoiesis of indeterminate potential (CHIP) are at increased risk of aging related health conditions and all-cause mortality, but whether CHIP affects risk of infection is much less clear. Using UK Biobank data, we revealed a positive association between CHIP and incident pneumonia in 438,421 individuals. We show that inflammation enhanced pneumonia risk, as CHIP carriers with a hypomorphic IL6 receptor polymorphism were protected. To better characterize the pathways of susceptibility, we challenged hematopoietic Tet Methylcytosine Dioxygenase 2-knockout (Tet2-/-) and floxed control mice (Tet2fl/fl) with Streptococcus pneumoniae. As with human CHIP carriers, Tet2-/- mice had hematopoietic abnormalities resulting in the expansion of inflammatory monocytes and neutrophils in peripheral blood. Yet, these cells were insufficient in defending against S. pneumoniae and resulted in increased pathology, impaired bacterial clearance, and higher mortality in Tet2-/- mice. We delineated the transcriptional landscape of Tet2-/- neutrophils and found that, while inflammation-related pathways were upregulated in Tet2-/- neutrophils, migration and motility pathways were compromised. Using live-imaging techniques, we demonstrated impairments in motility, pathogen uptake, and neutrophil extracellular trap (NET) formation by Tet2-/- neutrophils. Collectively, we show that CHIP is a risk factor for bacterial pneumonia related to innate immune impairments.
Keywords: Immunology; Infectious disease; Innate immunity.
Conflict of interest statement
Figures


Comment in
- CHIPing away at immunity: the role of clonal hematopoiesis of indeterminate potential in bacterial pneumonia
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
