

Impaired amygdala astrocytic signaling worsens neuropathic pain-associated neuronal functions and behaviors
- PMID: 38576475
- PMCID: PMC10991799
- DOI: 10.3389/fphar.2024.1368634
Impaired amygdala astrocytic signaling worsens neuropathic pain-associated neuronal functions and behaviors
Abstract
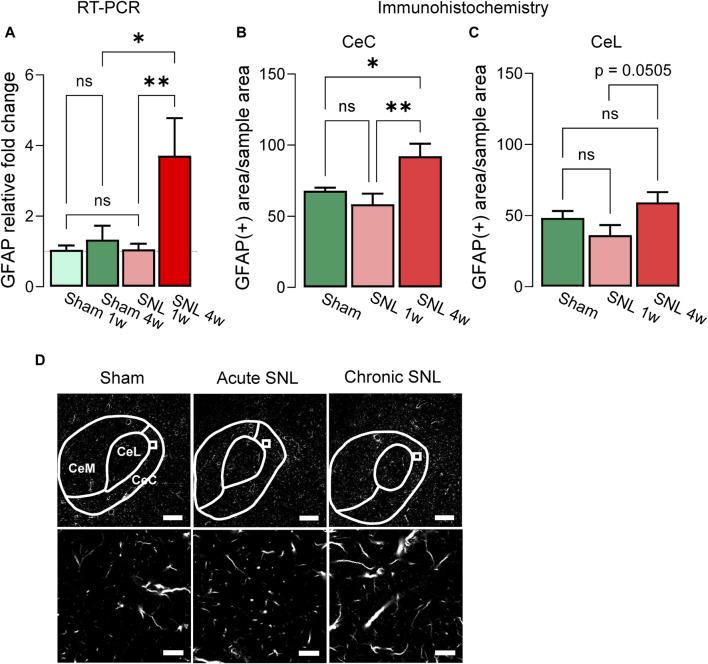
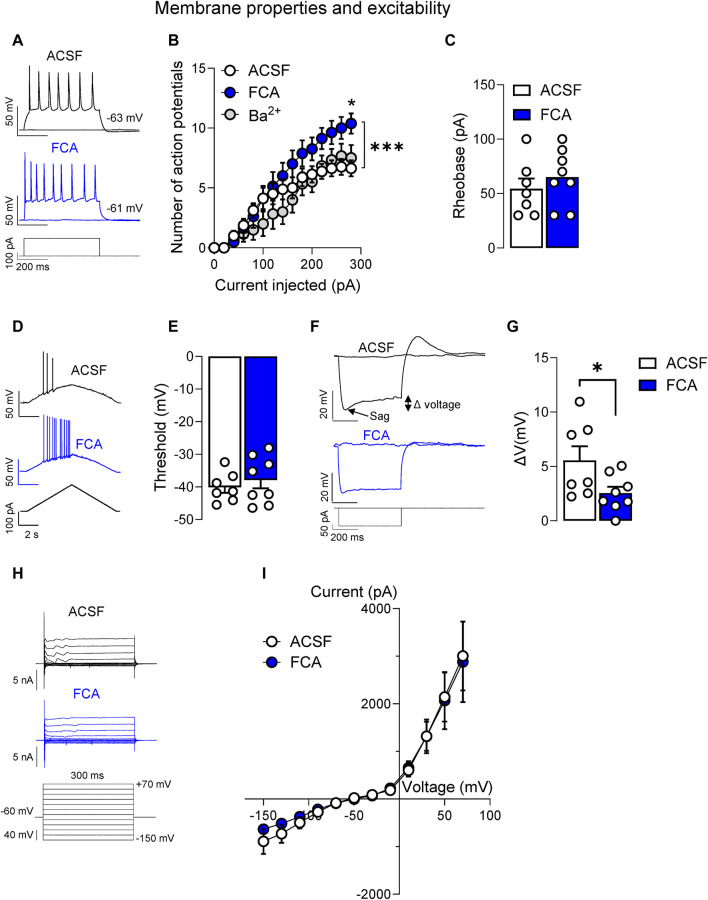
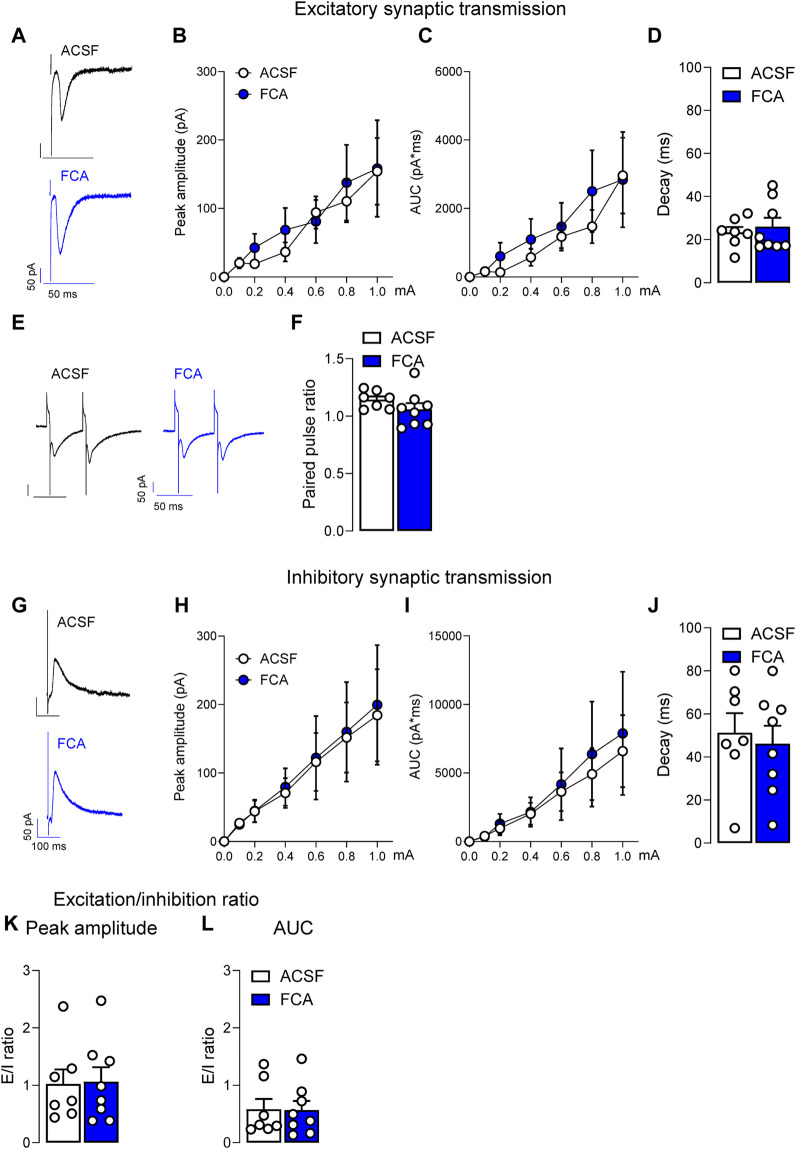
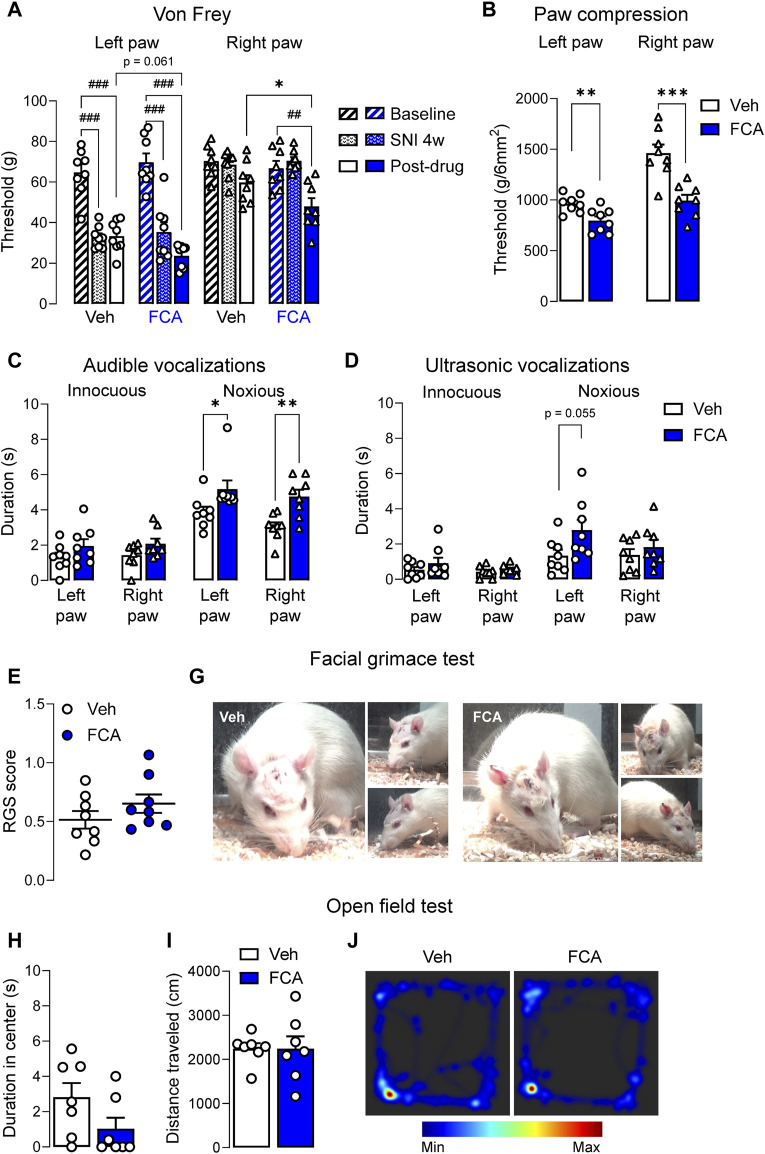
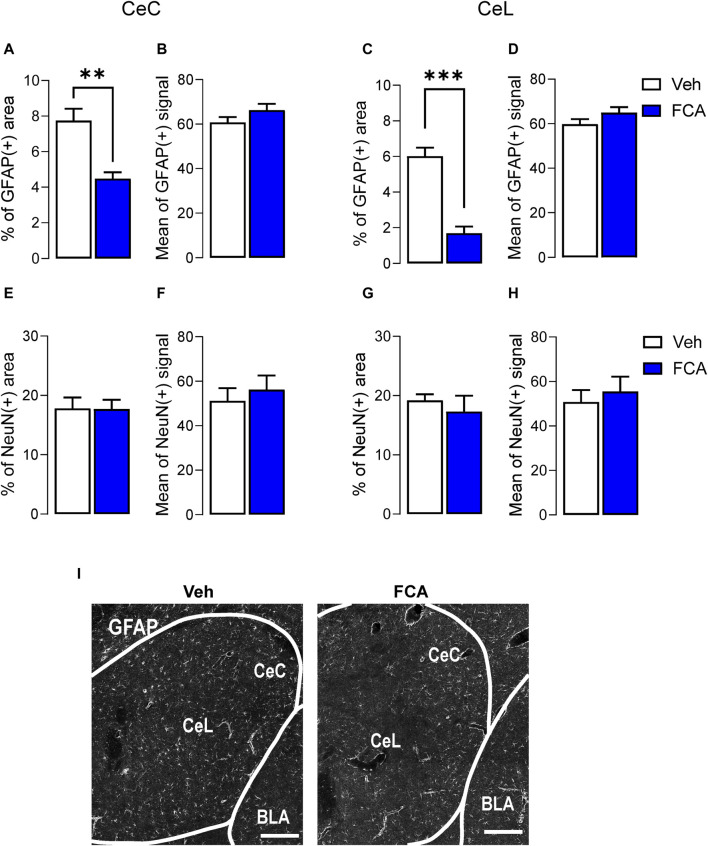
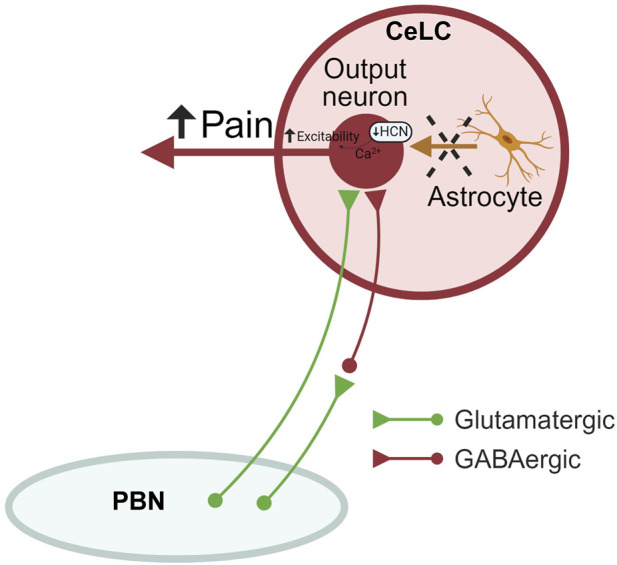
Introduction: Pain is a clinically relevant health care issue with limited therapeutic options, creating the need for new and improved analgesic strategies. The amygdala is a limbic brain region critically involved in the regulation of emotional-affective components of pain and in pain modulation. The central nucleus of amygdala (CeA) serves major output functions and receives nociceptive information via the external lateral parabrachial nucleus (PB). While amygdala neuroplasticity has been linked causally to pain behaviors, non-neuronal pain mechanisms in this region remain to be explored. As an essential part of the neuroimmune system, astrocytes that represent about 40-50% of glia cells within the central nervous system, are required for physiological neuronal functions, but their role in the amygdala remains to be determined for pain conditions. In this study, we measured time-specific astrocyte activation in the CeA in a neuropathic pain model (spinal nerve ligation, SNL) and assessed the effects of astrocyte inhibition on amygdala neuroplasticity and pain-like behaviors in the pain condition. Methods and Results: Glial fibrillary acidic protein (GFAP, astrocytic marker) immunoreactivity and mRNA expression were increased at the chronic (4 weeks post-SNL), but not acute (1 week post-SNL), stage of neuropathic pain. In order to determine the contribution of astrocytes to amygdala pain-mechanisms, we used fluorocitric acid (FCA), a selective inhibitor of astrocyte metabolism. Whole-cell patch-clamp recordings were performed from neurons in the laterocapsular division of the CeA (CeLC) obtained from chronic neuropathic rats. Pre-incubation of brain slices with FCA (100 µM, 1 h), increased excitability through altered hyperpolarization-activated current (Ih) functions, without significantly affecting synaptic responses at the PB-CeLC synapse. Intra-CeA injection of FCA (100 µM) had facilitatory effects on mechanical withdrawal thresholds (von Frey and paw pressure tests) and emotional-affective behaviors (evoked vocalizations), but not on facial grimace score and anxiety-like behaviors (open field test), in chronic neuropathic rats. Selective inhibition of astrocytes by FCA was confirmed with immunohistochemical analyses showing decreased astrocytic GFAP, but not NeuN, signal in the CeA. Discussion: Overall, these results suggest a complex modulation of amygdala pain functions by astrocytes and provide evidence for beneficial functions of astrocytes in CeA in chronic neuropathic pain.
Keywords: amygdala; astrocyte; behavior; electrophysiology; neuroimmune signaling; neuronal excitability; neuropathic pain; neurotransmission.
Copyright © 2024 Mazzitelli, Ponomareva, Presto, John and Neugebauer.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous