

TRPA5 encodes a thermosensitive ankyrin ion channel receptor in a triatomine insect
- PMID: 38577108
- PMCID: PMC10993193
- DOI: 10.1016/j.isci.2024.109541
TRPA5 encodes a thermosensitive ankyrin ion channel receptor in a triatomine insect
Abstract
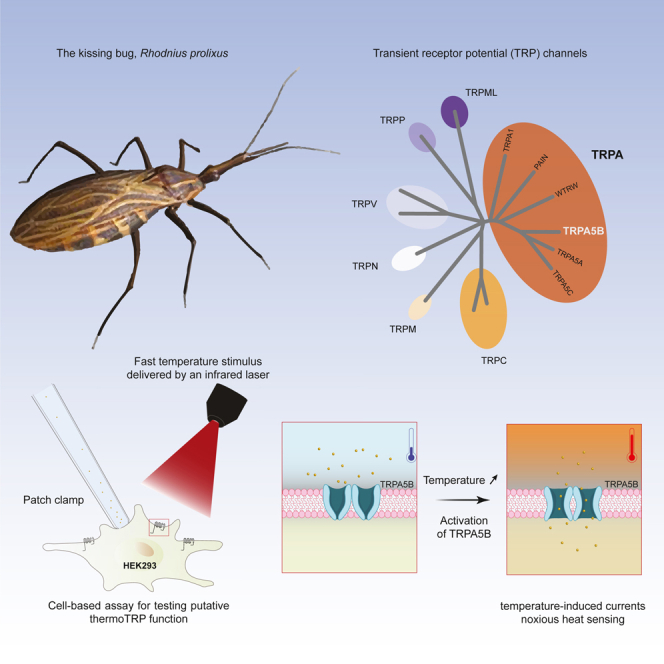
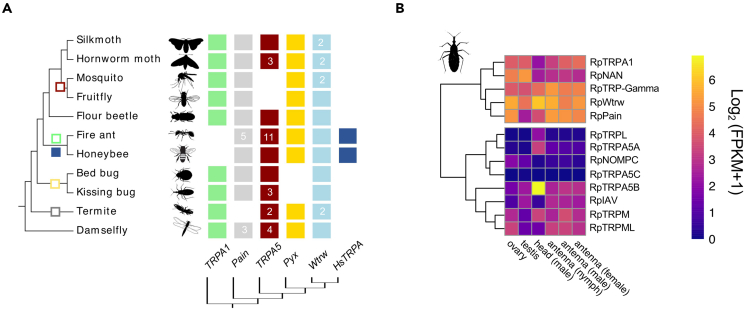
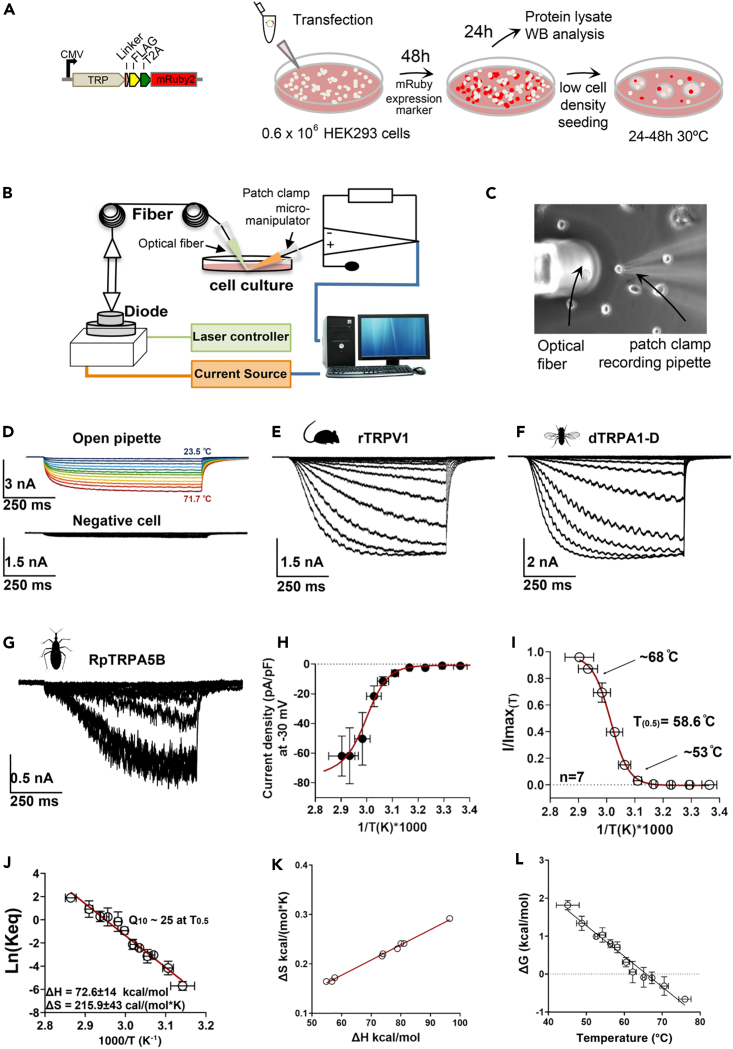
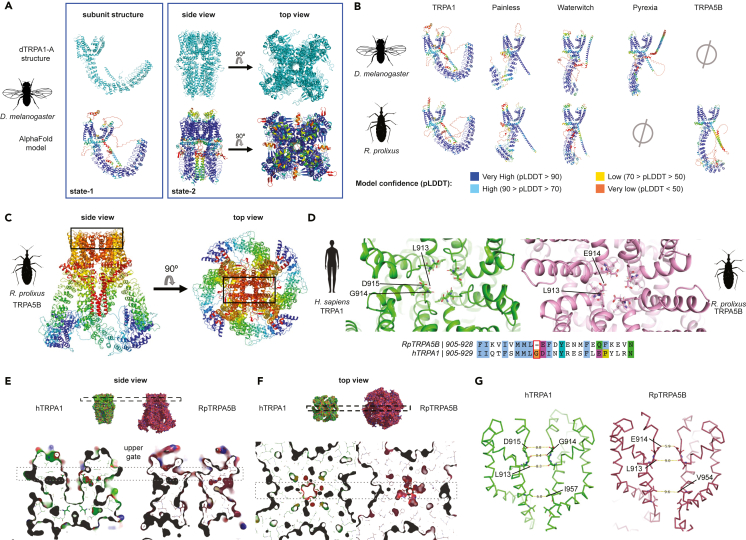
As ectotherms, insects need heat-sensitive receptors to monitor environmental temperatures and facilitate thermoregulation. We show that TRPA5, a class of ankyrin transient receptor potential (TRP) channels absent in dipteran genomes, may function as insect heat receptors. In the triatomine bug Rhodnius prolixus (order: Hemiptera), a vector of Chagas disease, the channel RpTRPA5B displays a uniquely high thermosensitivity, with biophysical determinants including a large channel activation enthalpy change (72 kcal/mol), a high temperature coefficient (Q10 = 25), and in vitro temperature-induced currents from 53°C to 68°C (T0.5 = 58.6°C), similar to noxious TRPV receptors in mammals. Monomeric and tetrameric ion channel structure predictions show reliable parallels with fruit fly dTRPA1, with structural uniqueness in ankyrin repeat domains, the channel selectivity filter, and potential TRP functional modulator regions. Overall, the finding of a member of TRPA5 as a temperature-activated receptor illustrates the diversity of insect molecular heat detectors.
Keywords: Biological sciences; Entomology; Molecular biology.
© 2024 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Thermosensation and the TRPV channel in Rhodnius prolixus.J Insect Physiol. 2015 Oct;81:145-56. doi: 10.1016/j.jinsphys.2015.07.014. Epub 2015 Jul 28. J Insect Physiol. 2015. PMID: 26225467
-
Trypanosoma cruzi, etiological agent of Chagas disease, is virulent to its triatomine vector Rhodnius prolixus in a temperature-dependent manner.PLoS Negl Trop Dis. 2015 Mar 20;9(3):e0003646. doi: 10.1371/journal.pntd.0003646. eCollection 2015 Mar. PLoS Negl Trop Dis. 2015. PMID: 25793495 Free PMC article.
-
Identification of a helix-turn-helix motif for high temperature dependence of vanilloid receptor TRPV2.J Physiol. 2021 Nov;599(21):4831-4844. doi: 10.1113/JP282073. Epub 2021 Oct 3. J Physiol. 2021. PMID: 34605028
-
Species-specific temperature sensitivity of TRPA1.Temperature (Austin). 2015 Feb 11;2(2):214-26. doi: 10.1080/23328940.2014.1000702. eCollection 2015 Apr-Jun. Temperature (Austin). 2015. PMID: 27227025 Free PMC article. Review.
-
Thermotaxis, circadian rhythms, and TRP channels in Drosophila.Temperature (Austin). 2015 Feb 11;2(2):227-43. doi: 10.1080/23328940.2015.1004972. eCollection 2015 Apr-Jun. Temperature (Austin). 2015. PMID: 27227026 Free PMC article. Review.
Cited by
-
Temperature-sensing riboceptors.RNA Biol. 2024 Jan;21(1):1-6. doi: 10.1080/15476286.2024.2379118. Epub 2024 Jul 17. RNA Biol. 2024. PMID: 39016038 Free PMC article.
-
Hot breath, quick exit: aphids flee mammalian heat via TRPA1.Sci China Life Sci. 2025 Jun 19. doi: 10.1007/s11427-025-2925-9. Online ahead of print. Sci China Life Sci. 2025. PMID: 40550996
-
Evolution of Sensory Receptors.Annu Rev Cell Dev Biol. 2024 Oct;40(1):353-379. doi: 10.1146/annurev-cellbio-120123-112853. Epub 2024 Sep 21. Annu Rev Cell Dev Biol. 2024. PMID: 38985841 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources
Research Materials
