

Structural constraints of pyocin S2 import through the ferripyoverdine receptor FpvAI
- PMID: 38577260
- PMCID: PMC10994204
- DOI: 10.1093/pnasnexus/pgae124
Structural constraints of pyocin S2 import through the ferripyoverdine receptor FpvAI
Abstract
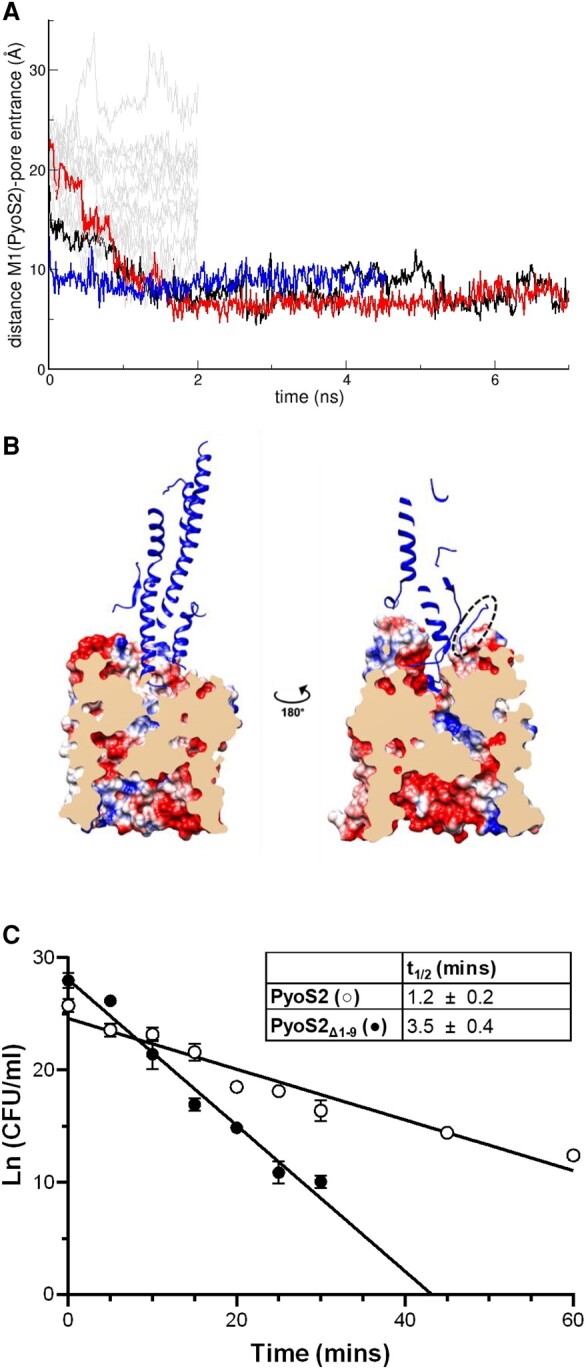
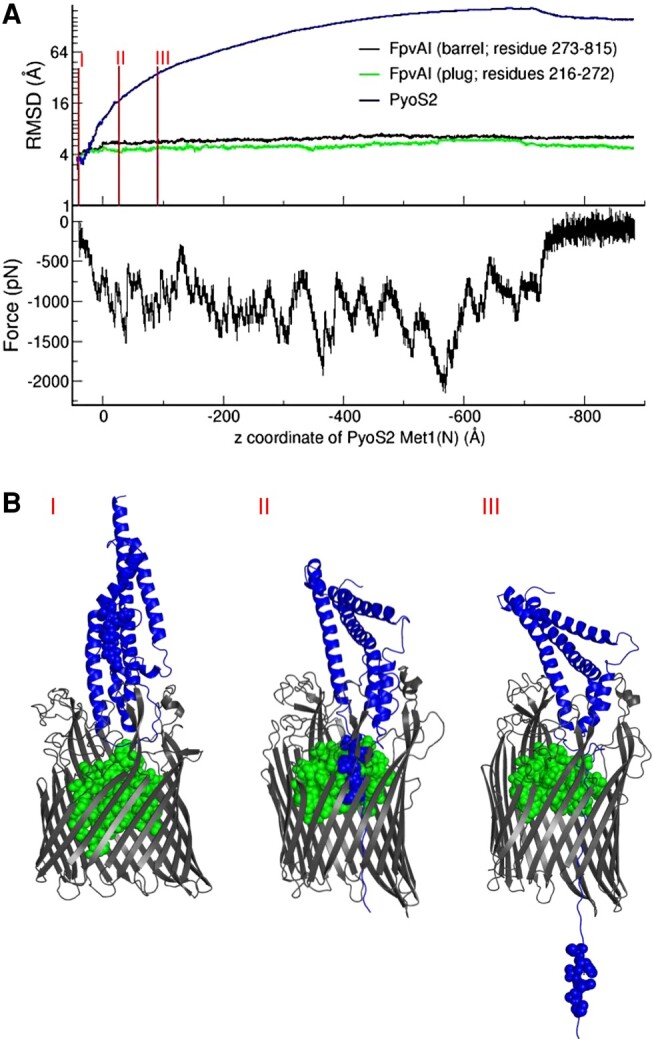
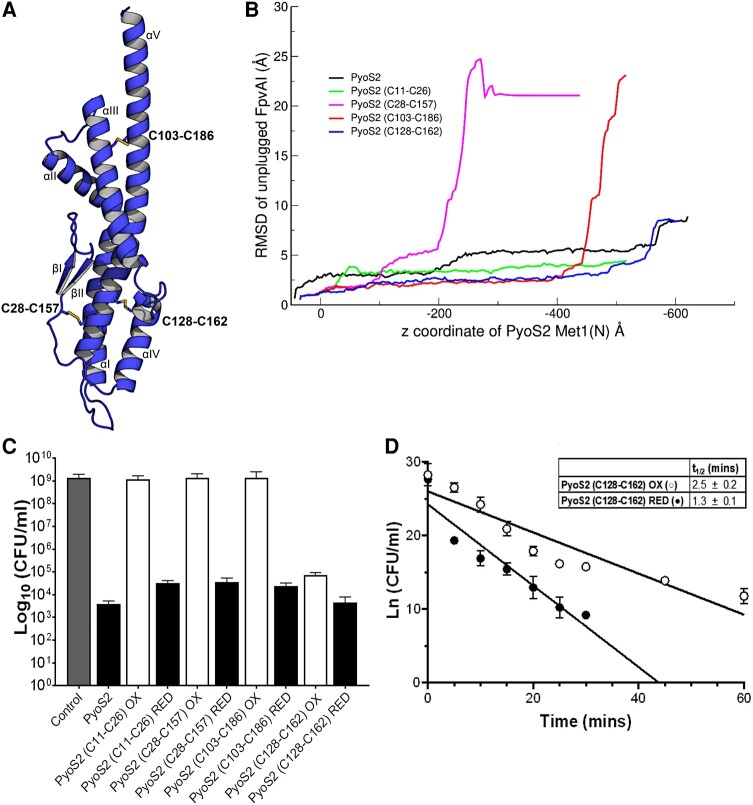
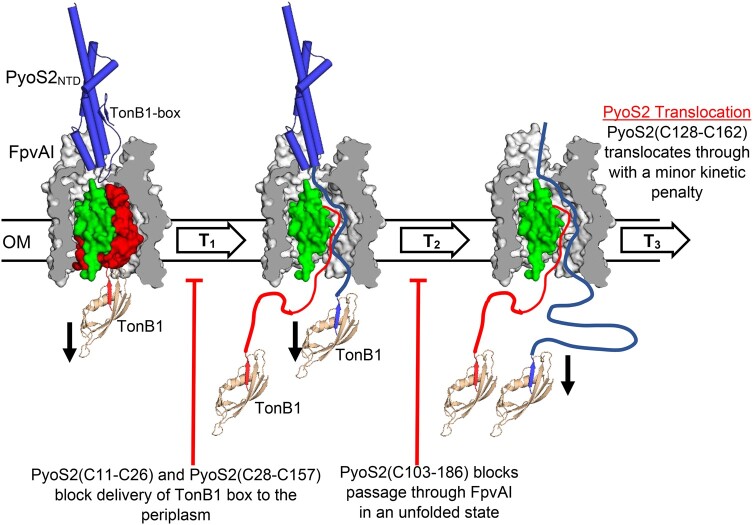
TonB-dependent transporters (TBDTs) mediate energized transport of essential nutrients into gram-negative bacteria. TBDTs are increasingly being exploited for the delivery of antibiotics to drug-resistant bacteria. While much is known about ground state complexes of TBDTs, few details have emerged about the transport process itself. In this study, we exploit bacteriocin parasitization of a TBDT to probe the mechanics of transport. Previous work has shown that the N-terminal domain of Pseudomonas aeruginosa-specific bacteriocin pyocin S2 (PyoS2NTD) is imported through the pyoverdine receptor FpvAI. PyoS2NTD transport follows the opening of a proton-motive force-dependent pore through FpvAI and the delivery of its own TonB box that engages TonB. We use molecular models and simulations to formulate a complete translocation pathway for PyoS2NTD that we validate using protein engineering and cytotoxicity measurements. We show that following partial removal of the FpvAI plug domain which occludes the channel, the pyocin's N-terminus enters the channel by electrostatic steering and ratchets to the periplasm. Application of force, mimicking that exerted by TonB, leads to unraveling of PyoS2NTD as it squeezes through the channel. Remarkably, while some parts of PyoS2NTD must unfold, complete unfolding is not required for transport, a result we confirmed by disulfide bond engineering. Moreover, the section of the FpvAI plug that remains embedded in the channel appears to serve as a buttress against which PyoS2NTD is pushed to destabilize the domain. Our study reveals the limits of structural deformation that accompanies import through a TBDT and the role the TBDT itself plays in accommodating transport.
© The Author(s) 2024. Published by Oxford University Press on behalf of National Academy of Sciences.
Figures

Similar articles
-
Exploitation of an iron transporter for bacterial protein antibiotic import.Proc Natl Acad Sci U S A. 2017 Nov 7;114(45):12051-12056. doi: 10.1073/pnas.1713741114. Epub 2017 Oct 25. Proc Natl Acad Sci U S A. 2017. PMID: 29078392 Free PMC article.
-
The soluble pyocins S2 and S4 from Pseudomonas aeruginosa bind to the same FpvAI receptor.Microbiologyopen. 2012 Sep;1(3):268-75. doi: 10.1002/mbo3.27. Epub 2012 Jun 22. Microbiologyopen. 2012. PMID: 23170226 Free PMC article.
-
TonB-dependent outer membrane transport: going for Baroque?Curr Opin Struct Biol. 2005 Aug;15(4):394-400. doi: 10.1016/j.sbi.2005.07.001. Curr Opin Struct Biol. 2005. PMID: 16039843 Review.
-
TonB-Dependent Transporters in Sphingomonads: Unraveling Their Distribution and Function in Environmental Adaptation.Microorganisms. 2020 Mar 3;8(3):359. doi: 10.3390/microorganisms8030359. Microorganisms. 2020. PMID: 32138166 Free PMC article. Review.
-
Energization of Outer Membrane Transport by the ExbB ExbD Molecular Motor.J Bacteriol. 2023 Jun 27;205(6):e0003523. doi: 10.1128/jb.00035-23. Epub 2023 May 23. J Bacteriol. 2023. PMID: 37219427 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
