

Genome-wide CRISPR screens identify novel regulators of wild-type and mutant p53 stability
- PMID: 38580884
- PMCID: PMC11148184
- DOI: 10.1038/s44320-024-00032-x
Genome-wide CRISPR screens identify novel regulators of wild-type and mutant p53 stability
Abstract
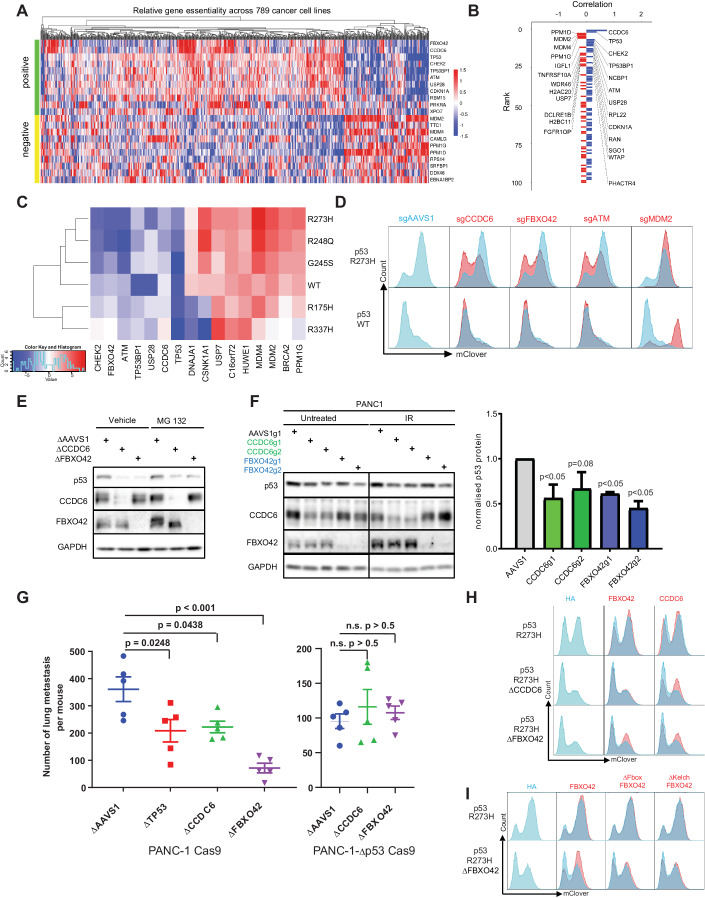
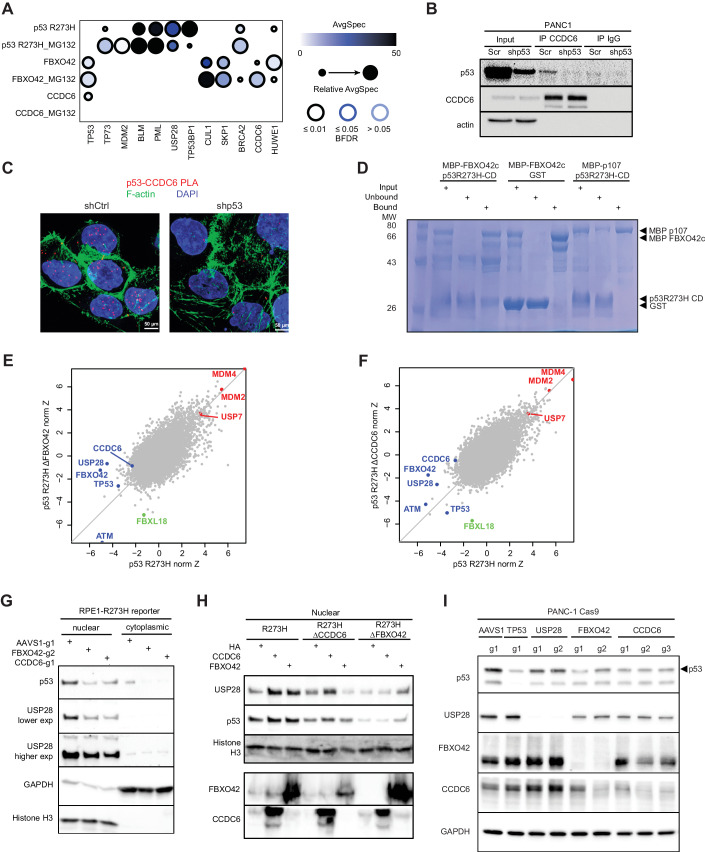
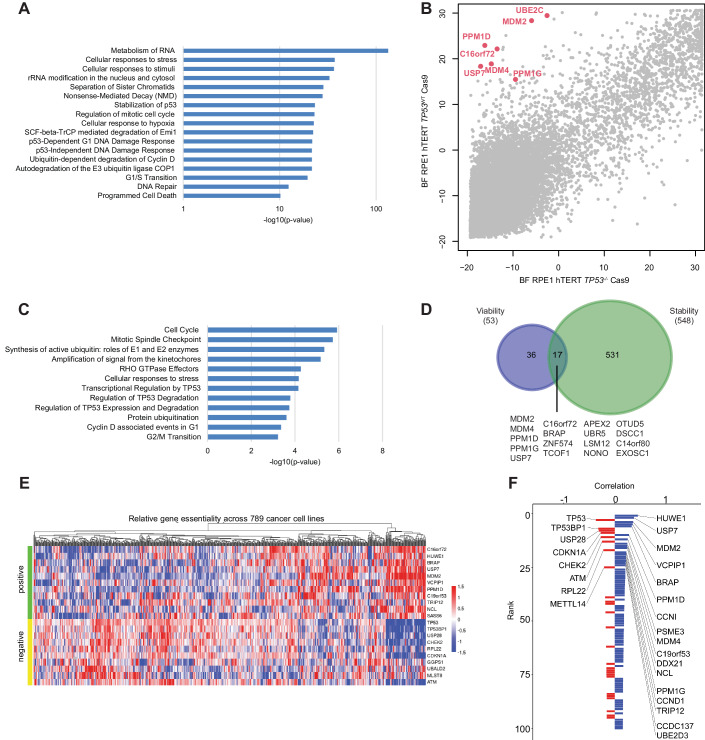
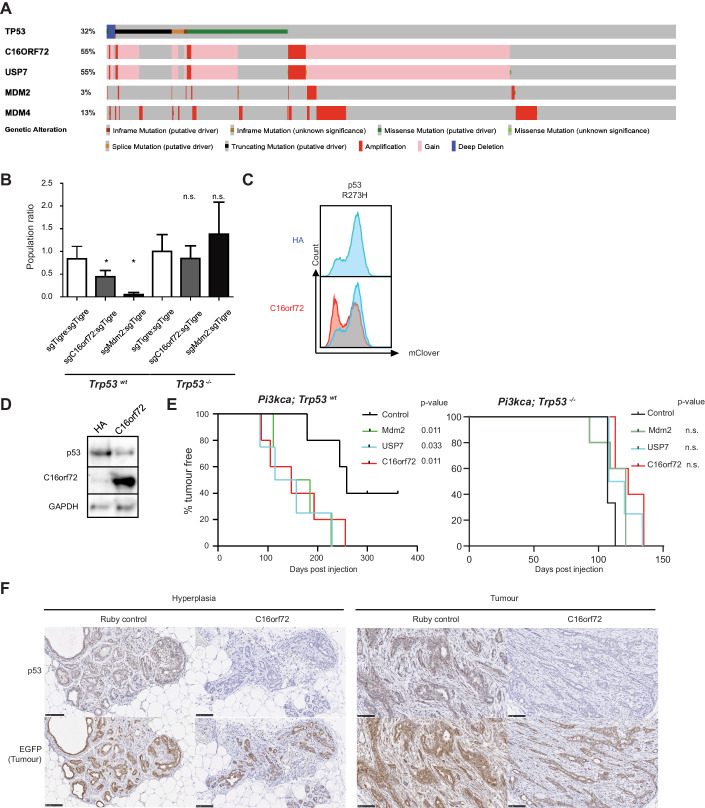
Tumor suppressor p53 (TP53) is frequently mutated in cancer, often resulting not only in loss of its tumor-suppressive function but also acquisition of dominant-negative and even oncogenic gain-of-function traits. While wild-type p53 levels are tightly regulated, mutants are typically stabilized in tumors, which is crucial for their oncogenic properties. Here, we systematically profiled the factors that regulate protein stability of wild-type and mutant p53 using marker-based genome-wide CRISPR screens. Most regulators of wild-type p53 also regulate p53 mutants, except for p53 R337H regulators, which are largely private to this mutant. Mechanistically, FBXO42 emerged as a positive regulator for a subset of p53 mutants, working with CCDC6 to control USP28-mediated mutant p53 stabilization. Additionally, C16orf72/HAPSTR1 negatively regulates both wild-type p53 and all tested mutants. C16orf72/HAPSTR1 is commonly amplified in breast cancer, and its overexpression reduces p53 levels in mouse mammary epithelium leading to accelerated breast cancer. This study offers a network perspective on p53 stability regulation, potentially guiding strategies to reinforce wild-type p53 or target mutant p53 in cancer.
Keywords: Breast Cancer; Fluorescence-based Stability Reporter; Genome-wide CRISPR Screening; Mutant p53; p53 Stability.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests. ACG is an editorial advisory board member. This has no bearing on the editorial consideration of this article for publication.
Figures


References
-
- Adams JR, Xu K, Liu JC, Agamez NM, Loch AJ, Wong RG, Wang W, Wright KL, Lane TF, Zacksenhaus E, et al. Cooperation between Pik3ca and p53 mutations in mouse mammary tumor formation. Cancer Res. 2011;71:2706–2717. - PubMed
-
- Amici DR, Ansel DJ, Metz KA, Smith RS, Phoumyvong CM, Gayatri S, Chamera T, Edwards SL, O’Hara BP, Srivastava S, Brockway S, Takagishi SR, Cho BK, Goo YA, Kelleher NL, Ben-Sahra I, Foltz DR, Li J, Mendillo ML (2022) C16orf72/HAPSTR1 is a molecular rheostat in an integrated network of stress response pathways. Proc Natl Acad Sci USA 119(27):e2111262119 - PMC - PubMed
MeSH terms
Substances
Grants and funding
- 384428/International Development Research Centre (IDRC)
- 705644/CCS | Canadian Cancer Society Research Institute (CCSRI)
- 157921/Canadian Government | Canadian Institutes of Health Research (CIHR)
- MSFSS 431649/Canadian Government | Canadian Institutes of Health Research (CIHR)
- MSFSS 431649/Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
