

This is a preprint.
Multifaceted roles of cohesin in regulating transcriptional loops
- PMID: 38585764
- PMCID: PMC10996690
- DOI: 10.1101/2024.03.25.586715
Multifaceted roles of cohesin in regulating transcriptional loops
Abstract
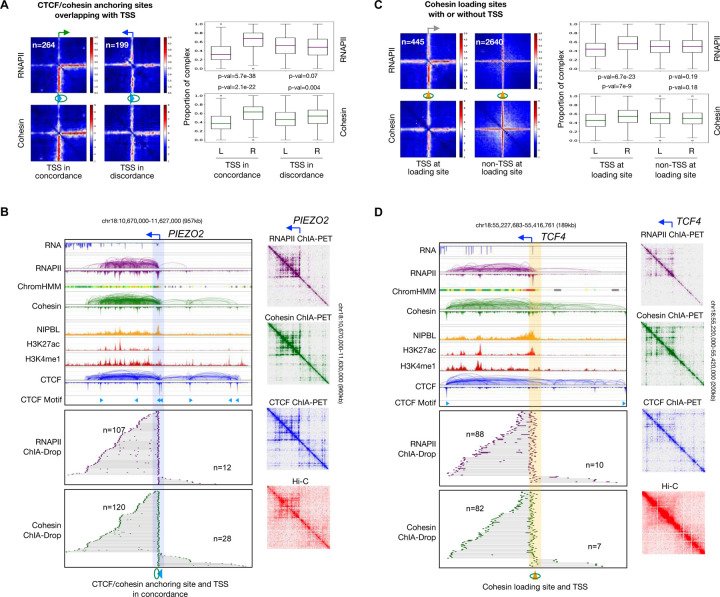
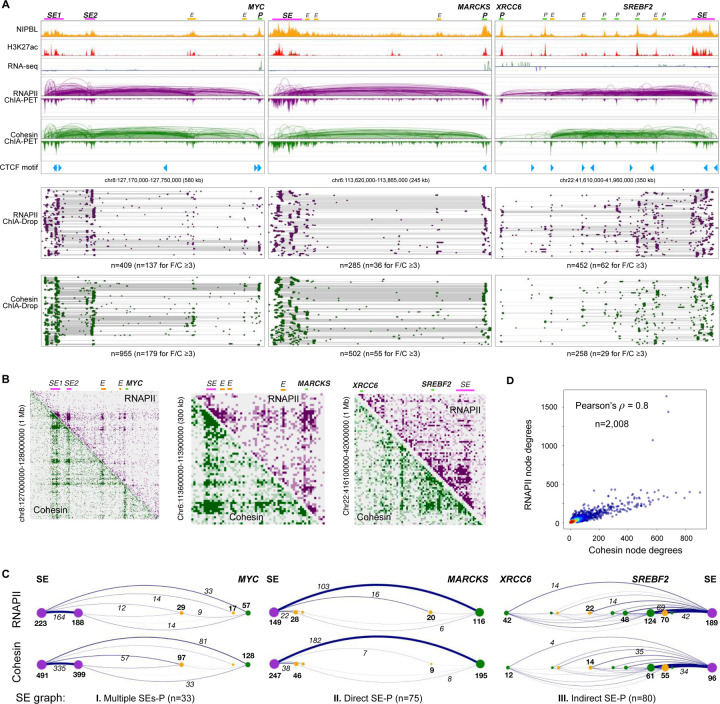
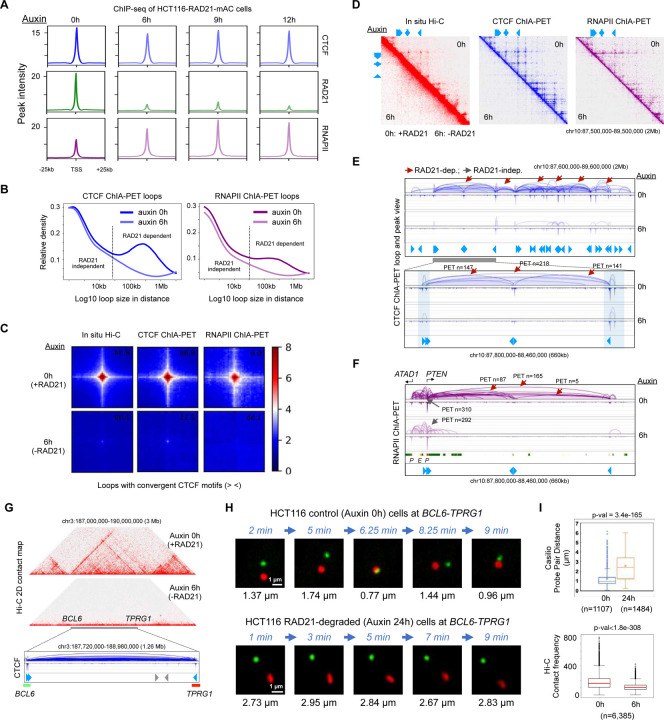
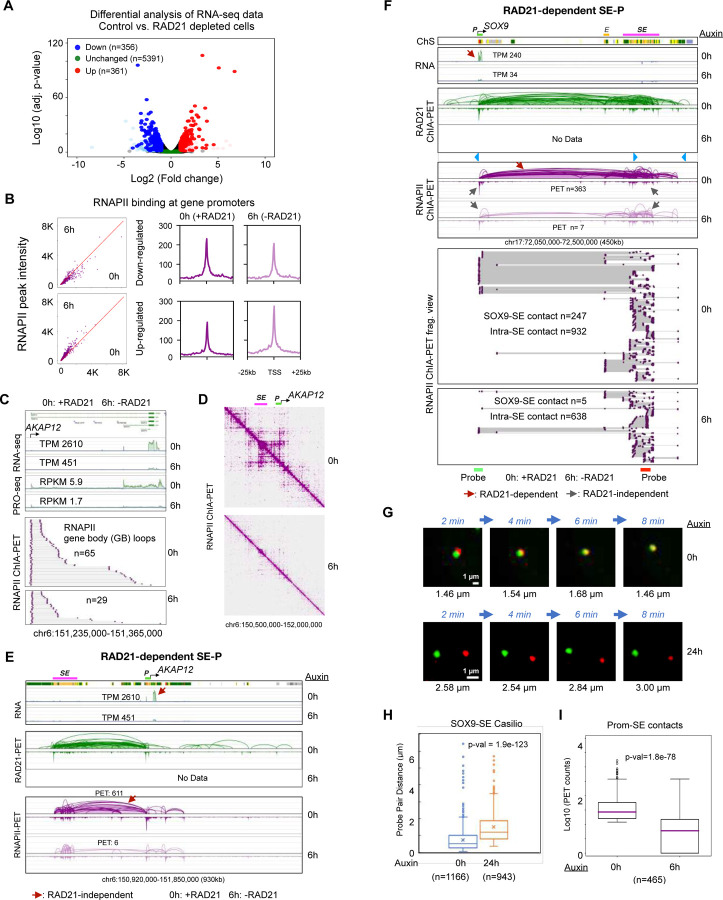
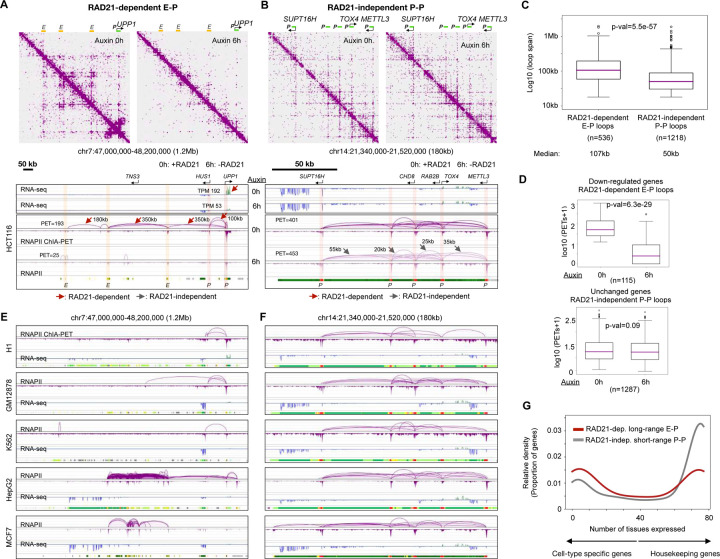
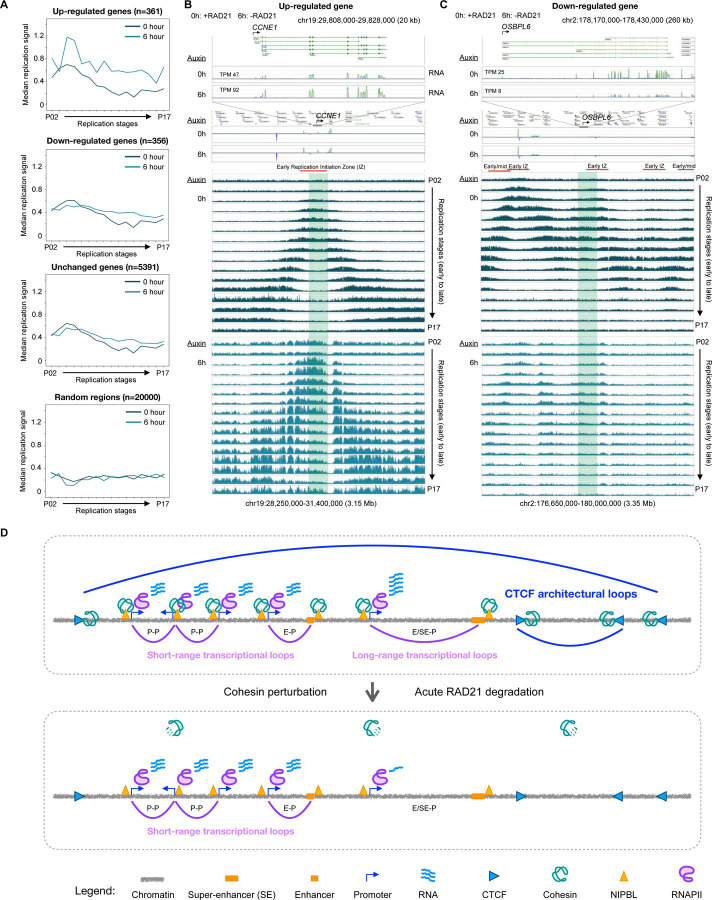
Cohesin is required for chromatin loop formation. However, its precise role in regulating gene transcription remains largely unknown. We investigated the relationship between cohesin and RNA Polymerase II (RNAPII) using single-molecule mapping and live-cell imaging methods in human cells. Cohesin-mediated transcriptional loops were highly correlated with those of RNAPII and followed the direction of gene transcription. Depleting RAD21, a subunit of cohesin, resulted in the loss of long-range (>100 kb) loops between distal (super-)enhancers and promoters of cell-type-specific genes. By contrast, the short-range (<50 kb) loops were insensitive to RAD21 depletion and connected genes that are mostly housekeeping. This result explains why only a small fraction of genes are affected by the loss of long-range chromatin interactions due to cohesin depletion. Remarkably, RAD21 depletion appeared to up-regulate genes located in early initiation zones (EIZ) of DNA replication, and the EIZ signals were amplified drastically without RAD21. Our results revealed new mechanistic insights of cohesin's multifaceted roles in establishing transcriptional loops, preserving long-range chromatin interactions for cell-specific genes, and maintaining timely order of DNA replication.
Keywords: 3D genome mapping; Cohesin; RNAPII; auxin-inducible degron (AID); chromatin loop formation; live-cell imaging; multiplex chromatin interaction; super-enhancers; transcription regulation.
Conflict of interest statement
DECLARATION OF INTERESTS The authors declare no competing interests.
Figures

References
-
- Allahyar A., Vermeulen C., Bouwman B.A., Krijger P.H., Verstegen M.J., Geeven G., van Kranenburg M., Pieterse M., Straver R., Haarhuis J.H. and Jalink K. (2018). Enhancer hubs and loop collisions identified from single-allele topologies. Nature genetics, 50(8), 1151–1160. - PubMed