

This is a preprint.
Synaptic-dependent developmental dysconnectivity in 22q11.2 deletion syndrome
- PMID: 38585897
- PMCID: PMC10996624
- DOI: 10.1101/2024.03.29.587339
Synaptic-dependent developmental dysconnectivity in 22q11.2 deletion syndrome
Update in
-
Synaptic-dependent developmental dysconnectivity in 22q11.2 deletion syndrome.Sci Adv. 2025 Mar 14;11(11):eadq2807. doi: 10.1126/sciadv.adq2807. Epub 2025 Mar 12. Sci Adv. 2025. PMID: 40073125 Free PMC article.
Abstract
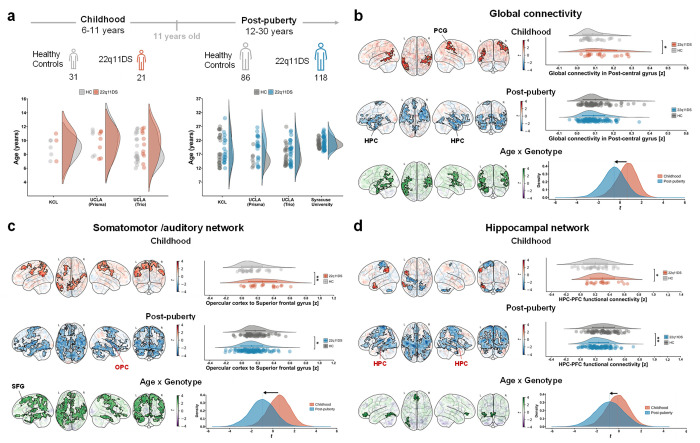
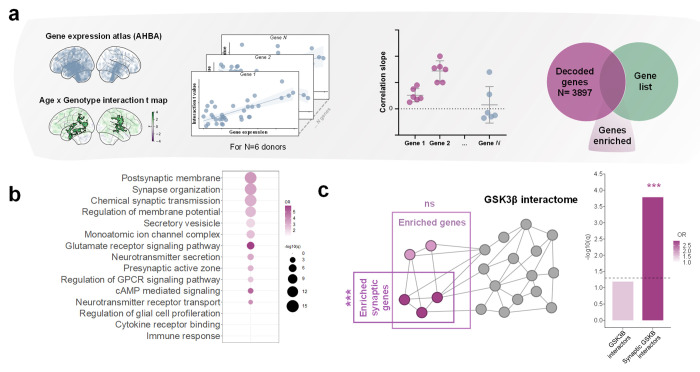
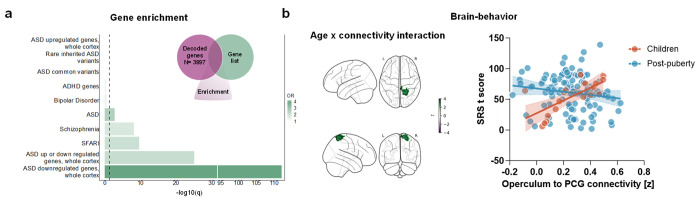
Chromosome 22q11.2 deletion is among the strongest known genetic risk factors for neuropsychiatric disorders, including autism and schizophrenia. Brain imaging studies have reported disrupted large-scale functional connectivity in people with 22q11 deletion syndrome (22q11DS). However, the significance and biological determinants of these functional alterations remain unclear. Here, we use a cross-species design to investigate the developmental trajectory and neural underpinnings of brain dysconnectivity in 22q11DS. We find that LgDel mice, an established mouse model of 22q11DS, exhibit age-specific patterns of functional MRI (fMRI) dysconnectivity, with widespread fMRI hyper-connectivity in juvenile mice reverting to focal hippocampal hypoconnectivity over puberty. These fMRI connectivity alterations are mirrored by co-occurring developmental alterations in dendritic spine density, and are both transiently normalized by developmental GSK3β inhibition, suggesting a synaptic origin for this phenomenon. Notably, analogous hyper- to hypoconnectivity reconfiguration occurs also in human 22q11DS, where it affects hippocampal and cortical regions spatially enriched for synaptic genes that interact with GSK3β, and autism-relevant transcripts. Functional dysconnectivity in somatomotor components of this network is predictive of age-dependent social alterations in 22q11.2 deletion carriers. Taken together, these findings suggest that synaptic-related mechanisms underlie developmentally mediated functional dysconnectivity in 22q11DS.
Figures


References
-
- Bullmore ET, Fornito A. Making connections: biological mechanisms of human brain (Dys) connectivity. Biological Psychiatry 93, 384–385 (2023). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources