

Transfer Learning Reveals Cancer-Associated Fibroblasts Are Associated with Epithelial-Mesenchymal Transition and Inflammation in Cancer Cells in Pancreatic Ductal Adenocarcinoma
- PMID: 38587552
- PMCID: PMC11065624
- DOI: 10.1158/0008-5472.CAN-23-1660
Transfer Learning Reveals Cancer-Associated Fibroblasts Are Associated with Epithelial-Mesenchymal Transition and Inflammation in Cancer Cells in Pancreatic Ductal Adenocarcinoma
Abstract
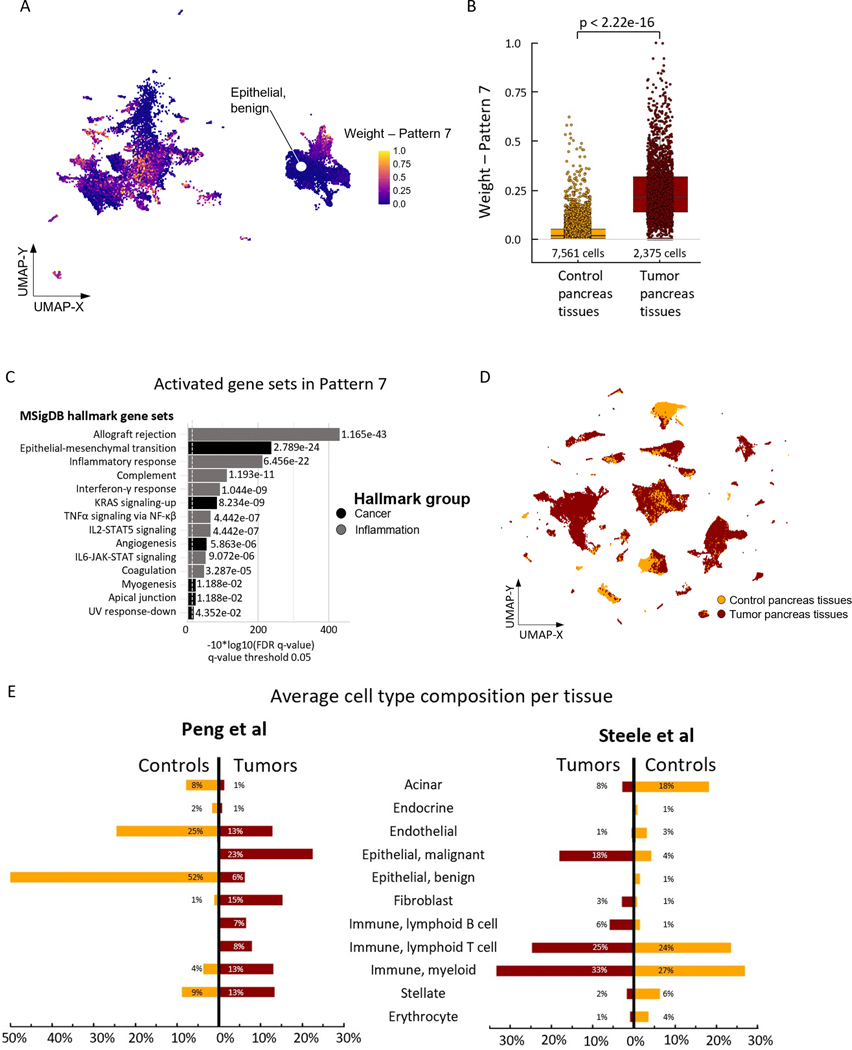
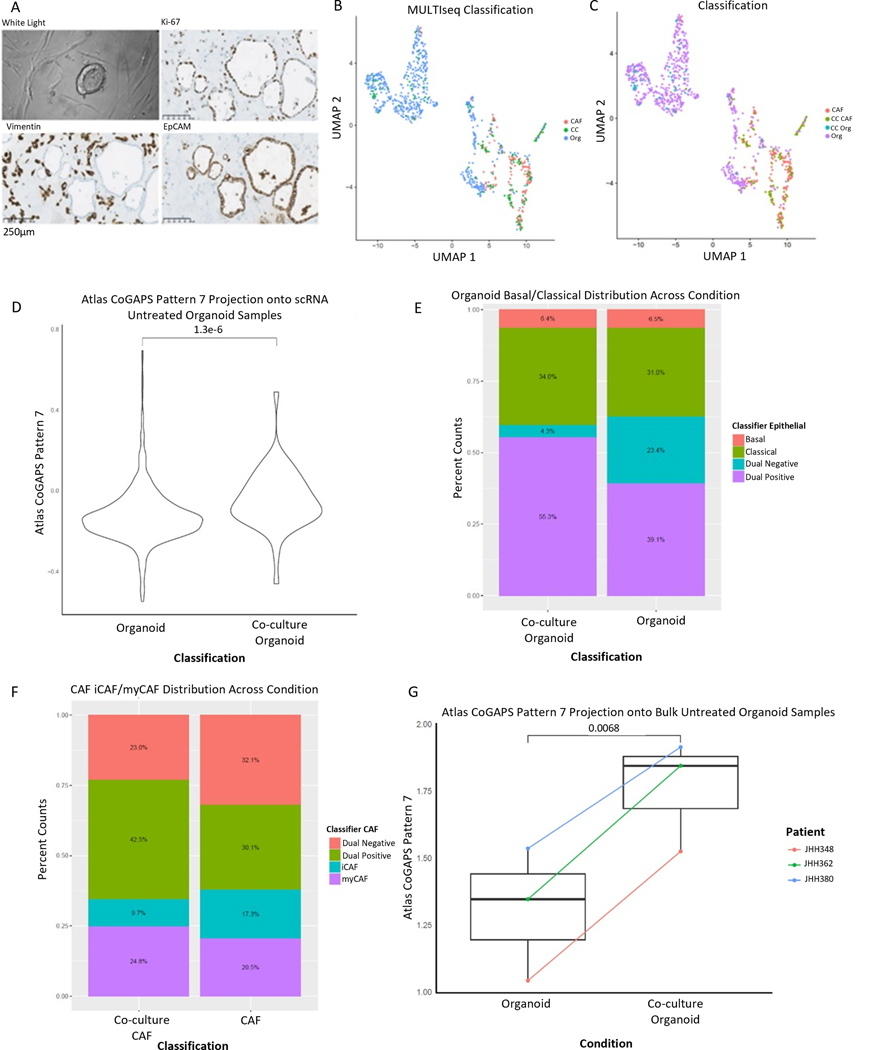
Pancreatic ductal adenocarcinoma (PDAC) is an aggressive malignancy characterized by an immunosuppressive tumor microenvironment enriched with cancer-associated fibroblasts (CAF). This study used a convergence approach to identify tumor cell and CAF interactions through the integration of single-cell data from human tumors with human organoid coculture experiments. Analysis of a comprehensive atlas of PDAC single-cell RNA sequencing data indicated that CAF density is associated with increased inflammation and epithelial-mesenchymal transition (EMT) in epithelial cells. Transfer learning using transcriptional data from patient-derived organoid and CAF cocultures provided in silico validation of CAF induction of inflammatory and EMT epithelial cell states. Further experimental validation in cocultures demonstrated integrin beta 1 (ITGB1) and vascular endothelial factor A (VEGFA) interactions with neuropilin-1 mediating CAF-epithelial cell cross-talk. Together, this study introduces transfer learning from human single-cell data to organoid coculture analyses for experimental validation of discoveries of cell-cell cross-talk and identifies fibroblast-mediated regulation of EMT and inflammation.
Significance: Adaptation of transfer learning to relate human single-cell RNA sequencing data to organoid-CAF cocultures facilitates discovery of human pancreatic cancer intercellular interactions and uncovers cross-talk between CAFs and tumor cells through VEGFA and ITGB1.
©2024 American Association for Cancer Research.
Conflict of interest statement
T.T.S. is the CEO and co-owner of Healthfund Finland and reports consultation fees from Boehringer Ingelheim Finland and Amgen. E.M.J is a paid consultant for Adaptive Biotech, Achilles, DragonFly, Candel Therapeutics, Genocea, and Roche. She receives funding from Lustgarten Foundation and Bristol Myer Squibb. She is the Chief Medical Advisor for Lustgarten and SAB advisor to the Parker Institute for Cancer Immunotherapy (PICI) and for the C3 Cancer Institute. She is a founding member of Abmeta. E.J.F is on the SAB for Resistance Biology, Consultant for Mestag Therapeutics and Merck. D.T.T. has received consulting fees from ROME Therapeutics, Tekla Capital, Ikena Oncology, Foundation Medicine, Inc., NanoString Technologies, and Pfizer that are not related to this work. D.T.T. is a founder and has equity in ROME Therapeutics, PanTher Therapeutics and TellBio, Inc., which is not related to this work. D.T.T. receives research support from ACD-Biotechne, PureTech Health LLC, and Ribon Therapeutics, which was not used in this work. D.T.T.’s interests were reviewed and are managed by Massachusetts General Hospital and Mass General Brigham in accordance with their conflict of interest policies. L.Z. reports personal fees from Biosion, Alphamab, NovaRock, Xilio, Ambrx, Novagenesis, and Snow Lake Capitals; and other support from Alphamab and Mingruizhiyao outside the submitted work. A.C.K. reports support from Vescor Therapeutics, Rafael Pharma, and AbbVie outside the submitted work; in addition, A.C.K. has a patent for targeting alanine transport pending, a patent for KRAS-regulated metabolic pathways issued, a patent for targeting GOT1 as a therapeutic approach issued, and a patent for autophagy control of iron metabolism issued. D.P.R. reports personal fees and other support from MPM, other support from Boehringer Ingelheim and Exact Sciences, and personal fees from UpToDate and McGraw Hill outside the submitted work. J.W.Z. reports grant funding from Genentch outside the submitted work.
Competing interests:
All other authors declare no potential conflict of interest.
Figures




References
-
- Peran I, Madhavan S, Byers SW, McCoy MD. Curation of the Pancreatic Ductal Adenocarcinoma Subset of the Cancer Genome Atlas Is Essential for Accurate Conclusions about Survival-Related Molecular Mechanisms. Clin Cancer Res. 2018. Aug 15;24(16):3813–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
