

Developmental trajectories of GABAergic cortical interneurons are sequentially modulated by dynamic FoxG1 expression levels
- PMID: 38588430
- PMCID: PMC11032493
- DOI: 10.1073/pnas.2317783121
Developmental trajectories of GABAergic cortical interneurons are sequentially modulated by dynamic FoxG1 expression levels
Abstract
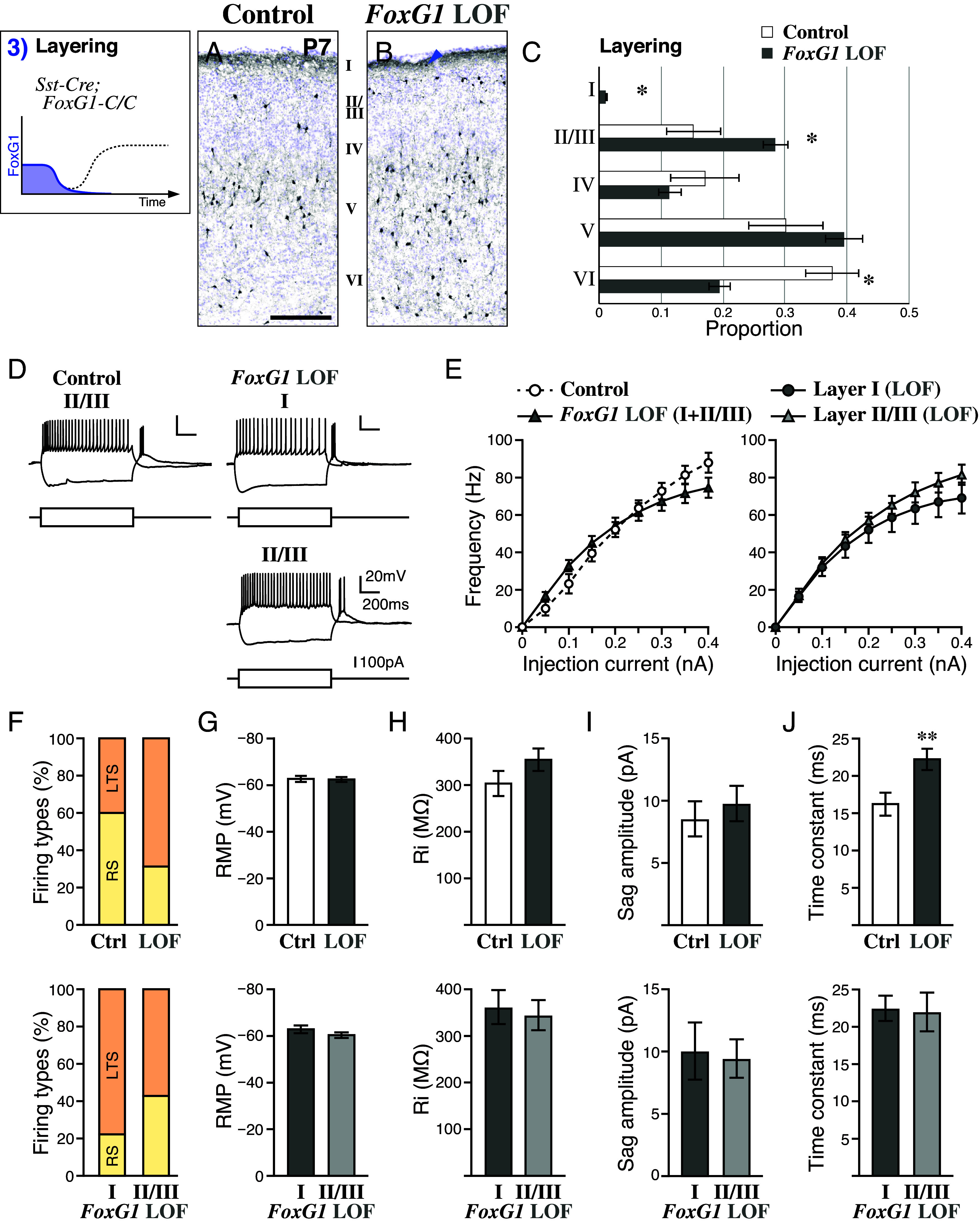
GABAergic inhibitory interneurons, originating from the embryonic ventral forebrain territories, traverse a convoluted migratory path to reach the neocortex. These interneuron precursors undergo sequential phases of tangential and radial migration before settling into specific laminae during differentiation. Here, we show that the developmental trajectory of FoxG1 expression is dynamically controlled in these interneuron precursors at critical junctures of migration. By utilizing mouse genetic strategies, we elucidate the pivotal role of precise changes in FoxG1 expression levels during interneuron specification and migration. Our findings underscore the gene dosage-dependent function of FoxG1, aligning with clinical observations of FOXG1 haploinsufficiency and duplication in syndromic forms of autism spectrum disorders. In conclusion, our results reveal the finely tuned developmental clock governing cortical interneuron development, driven by temporal dynamics and the dose-dependent actions of FoxG1.
Keywords: cortex; development; gene-dosage; inhibitory neuron; interneuron.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures





References
-
- Ma T., et al. , Subcortical origins of human and monkey neocortical interneurons. Nat. Neurosci. 16, 1588–1597 (2013). - PubMed
-
- Anderson S. A., Eisenstat D. D., Shi L., Rubenstein J. L., Interneuron migration from basal forebrain to neocortex: Dependence on Dlx genes. Science 278, 474–476 (1997). - PubMed
-
- Rakic P., Neurons in rhesus monkey visual cortex: Systematic relation between time of origin and eventual disposition. Science 183, 425–427 (1974). - PubMed
-
- Angevine J. B. J., Sidman R. L., Autoradiographic study of cell migration during histogenesis of cerebral cortex in the mouse. Nature 192, 766–768 (1961). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
