

EphB2 Receptor Promotes Dermal Fibrosis in Systemic Sclerosis
- PMID: 38589317
- PMCID: PMC11288787
- DOI: 10.1002/art.42858
EphB2 Receptor Promotes Dermal Fibrosis in Systemic Sclerosis
Abstract
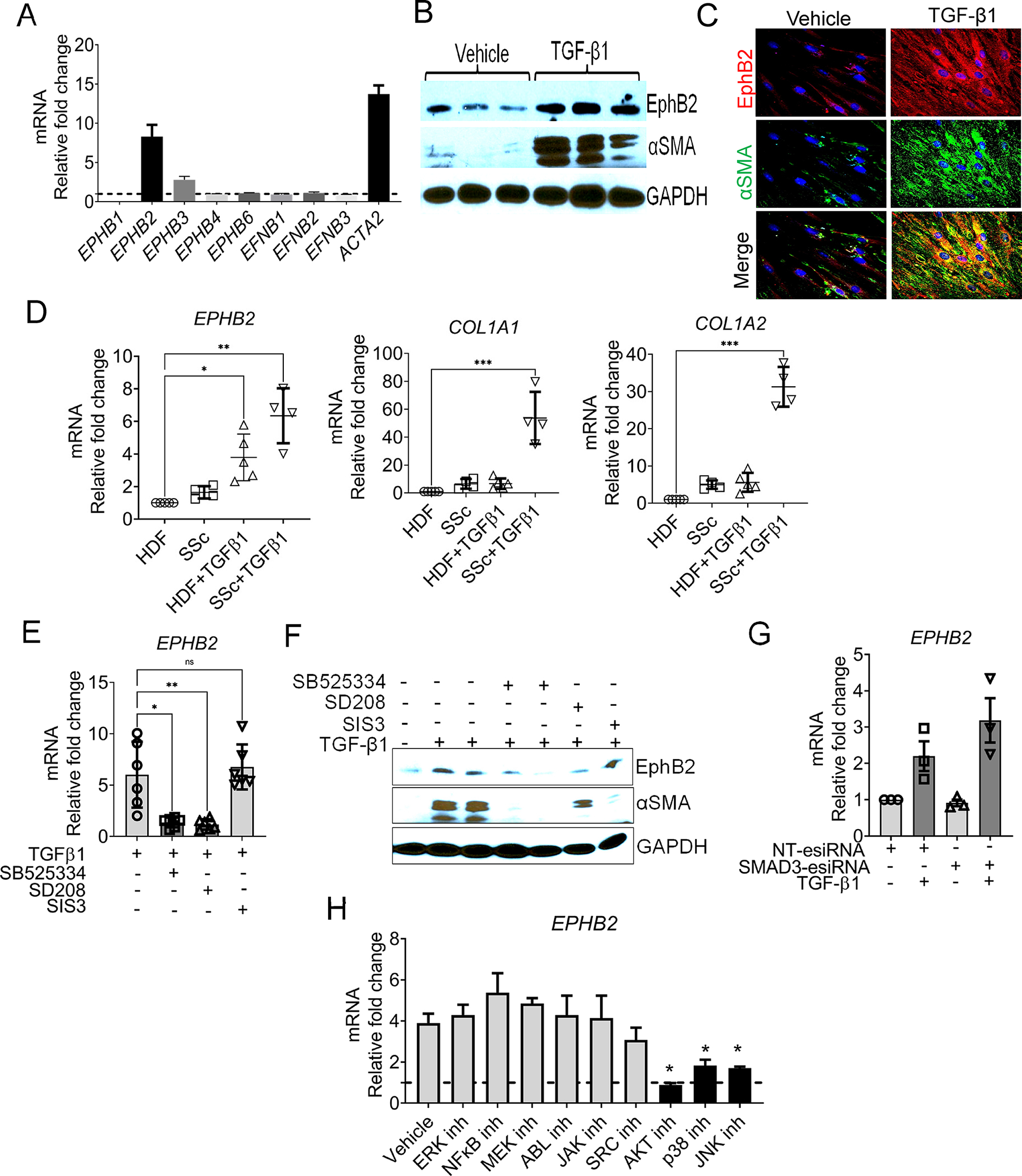
Objective: Erythropoietin-producing hepatocellular (Eph)/Ephrin cell-cell signaling is emerging as a key player in tissue fibrogenesis. The aim of this study was to test the hypothesis that the receptor tyrosine kinase EphB2 mediates dermal fibrosis in systemic sclerosis (SSc).
Methods: We assessed normal and SSc human skin biopsies for EphB2 expression. The in vivo role of EphB2 in skin fibrosis was investigated by subjecting EphB2-knockout mice to both bleomycin-induced and tight skin (Tsk1/+) genetic mouse models of skin fibrosis. EphB2 kinase-dead and overactive point mutant mice were used to evaluate the role of EphB2 forward signaling in bleomycin-induced dermal fibrosis. In vitro studies were performed on dermal fibroblasts from patients with SSc and healthy controls, which was followed by in vivo analysis of fibroblast-specific Ephb2-deficient mice.
Results: Expression of EphB2 is up-regulated in SSc skin tissue and explanted SSc dermal fibroblasts compared with healthy controls. EphB2 expression is elevated in two animal models of dermal fibrosis. In mice, EphB2 drives dermal fibrosis in both the bleomycin and the Tsk1/+ models of skin fibrosis. EphB2 forward signaling is a critical mediator of dermal fibrosis. Transforming growth factor-β (TGF-β) cytokines up-regulate EphB2 in dermal fibroblasts via noncanonical TGF-β/mother against decapentaplegic signaling, and silencing EPHB2 in human dermal fibroblasts is sufficient to dampen TGF-β-induced fibroblast-to-myofibroblast differentiation. Moreover, mice with fibroblast-specific deletion of EphB2 showed impaired fibroblast-to-myofibroblast differentiation and reduced skin fibrosis upon bleomycin challenge.
Conclusion: Our data implicate TGF-β regulation of EphB2 overexpression and kinase-mediated forward signaling in the development of dermal fibrosis in SSc. EphB2 thus represents a potential new therapeutic target for SSc.
© 2024 The Authors. Arthritis & Rheumatology published by Wiley Periodicals LLC on behalf of American College of Rheumatology.
Conflict of interest statement
Figures





References
-
- Stern EP, Denton CP. The Pathogenesis of Systemic Sclerosis. Rheumatic Disease Clinics of North America 2015;41:367–382. - PubMed
-
- Distler O, Highland KB, Gahlemann M, Azuma A, Fischer A, Mayes MD, Raghu G, et al. Nintedanib for Systemic Sclerosis–Associated Interstitial Lung Disease. New England Journal of Medicine 2019;380:2518–2528. - PubMed
-
- Kuwana M, Azuma A. Nintedanib: New indication for systemic sclerosis-associated interstitial lung disease. Modern Rheumatology 2019:1–7. - PubMed
-
- Denton CP. Advances in pathogenesis and treatment of systemic sclerosis. Clinical Medicine 2015;15:s58–s63. - PubMed
-
- Ide M, Jinnin M, Tomizawa Y, Wang Z, Kajihara I, Fukushima S, Hashizume Y, et al. Transforming growth factor β-inhibitor Repsox downregulates collagen expression of scleroderma dermal fibroblasts and prevents bleomycin-induced mice skin fibrosis. Experimental Dermatology:n/a-n/a. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
