

Lifelong absence of microglia alters hippocampal glutamatergic networks but not synapse and spine density
- PMID: 38589666
- PMCID: PMC11094096
- DOI: 10.1038/s44319-024-00130-9
Lifelong absence of microglia alters hippocampal glutamatergic networks but not synapse and spine density
Abstract
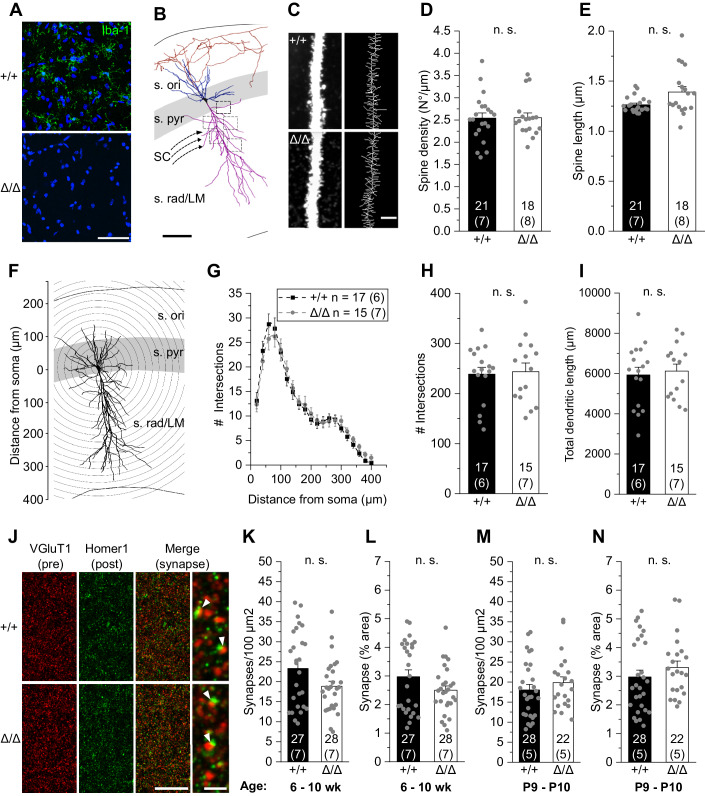
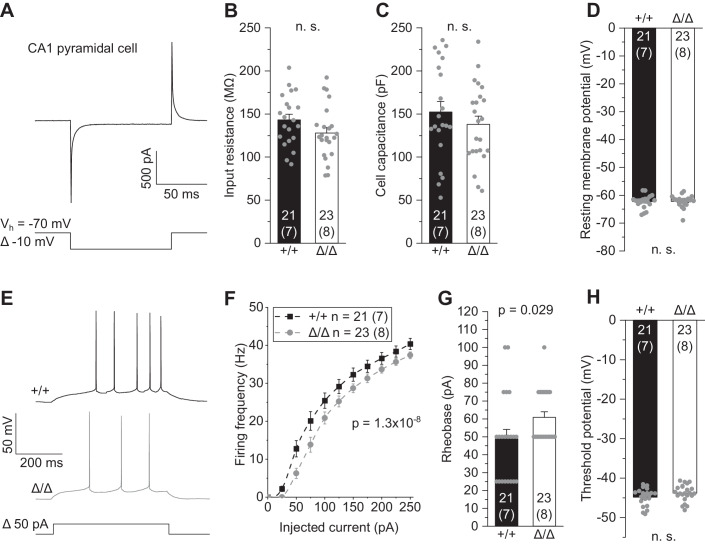
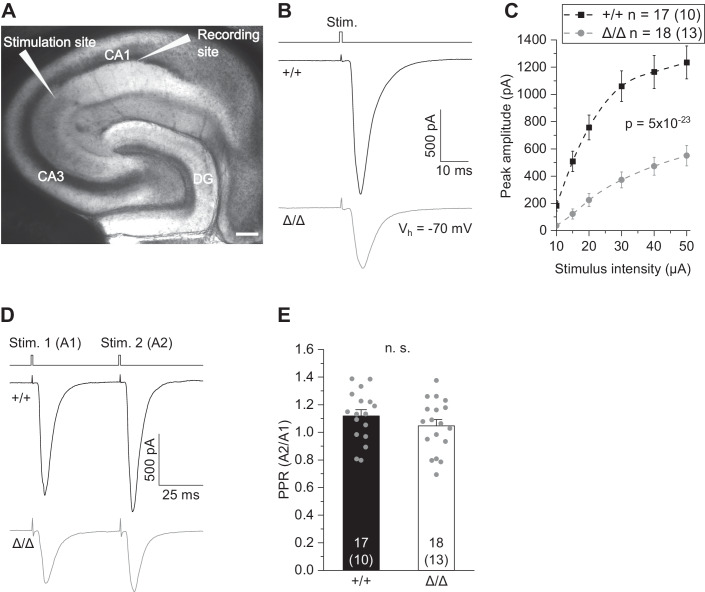
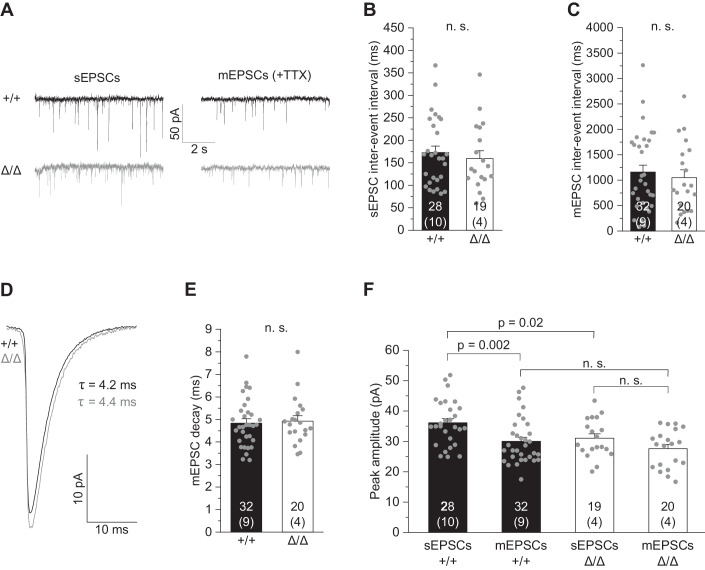
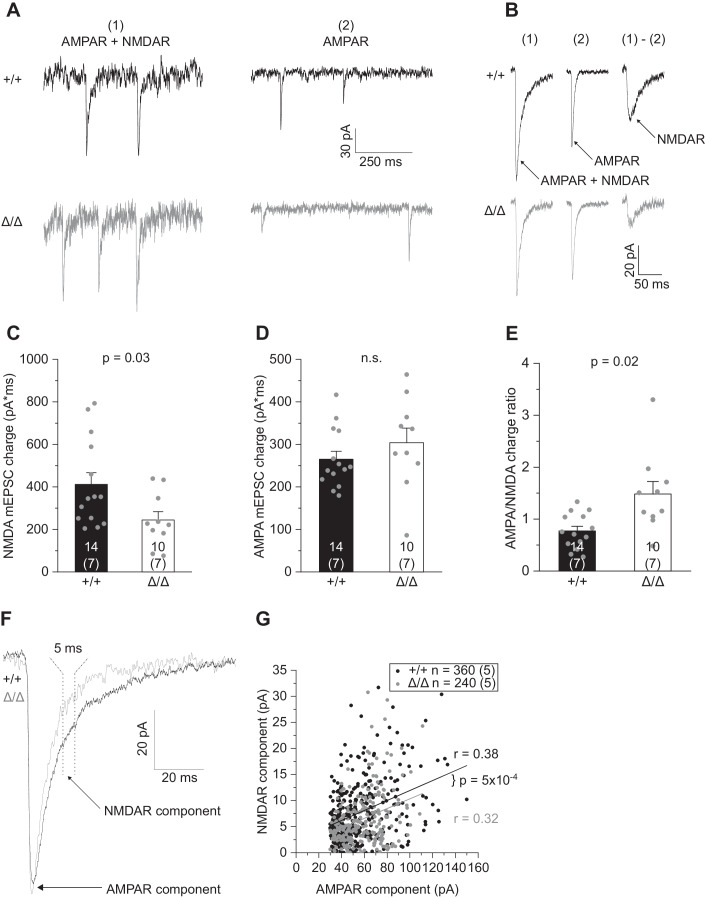
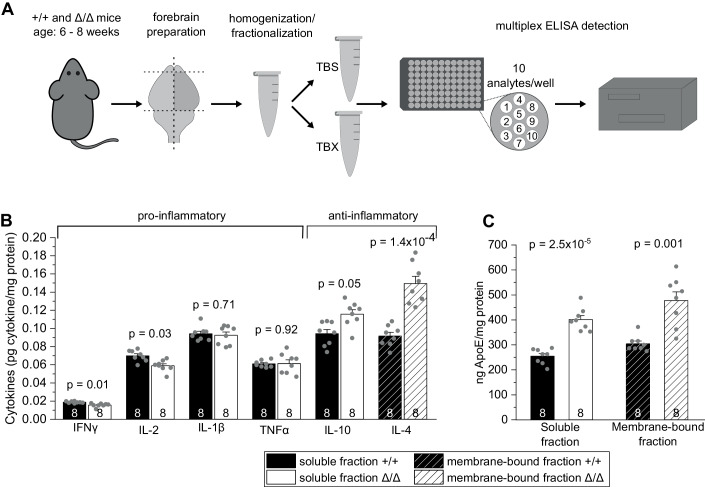
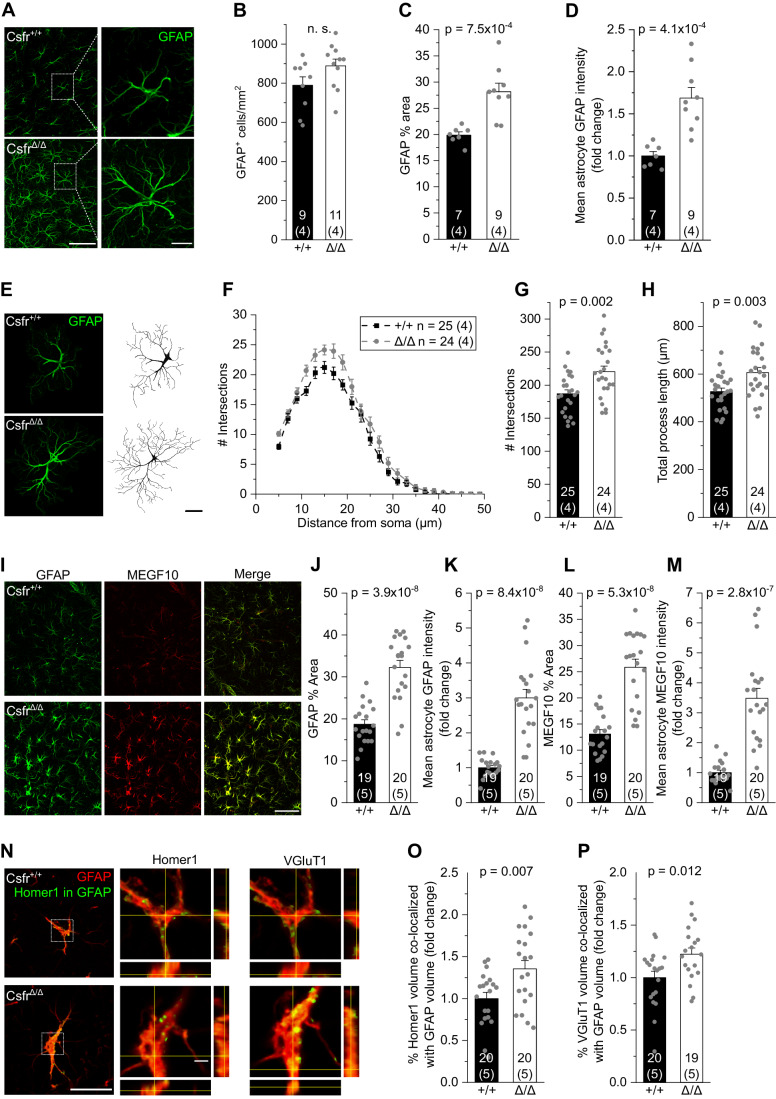
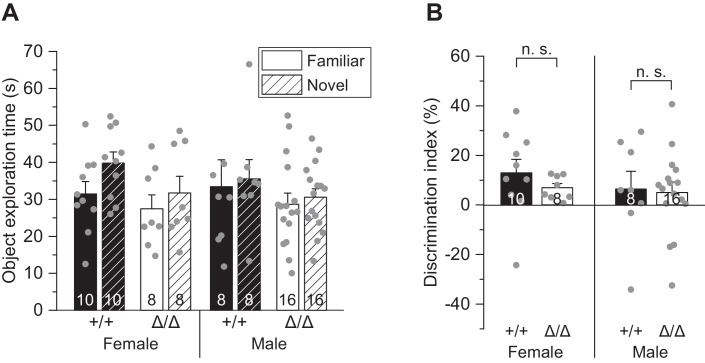
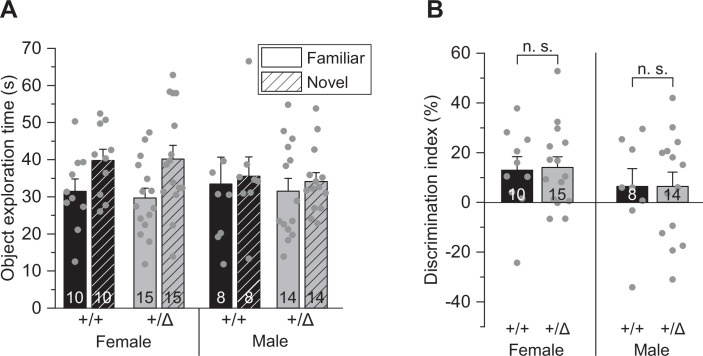
Microglia sculpt developing neural circuits by eliminating excess synapses in a process called synaptic pruning, by removing apoptotic neurons, and by promoting neuronal survival. To elucidate the role of microglia during embryonic and postnatal brain development, we used a mouse model deficient in microglia throughout life by deletion of the fms-intronic regulatory element (FIRE) in the Csf1r locus. Surprisingly, young adult Csf1rΔFIRE/ΔFIRE mice display no changes in excitatory and inhibitory synapse number and spine density of CA1 hippocampal neurons compared with Csf1r+/+ littermates. However, CA1 neurons are less excitable, receive less CA3 excitatory input and show altered synaptic properties, but this does not affect novel object recognition. Cytokine profiling indicates an anti-inflammatory state along with increases in ApoE levels and reactive astrocytes containing synaptic markers in Csf1rΔFIRE/ΔFIRE mice. Notably, these changes in Csf1rΔFIRE/ΔFIRE mice closely resemble the effects of acute microglial depletion in adult mice after normal development. Our findings suggest that microglia are not mandatory for synaptic pruning, and that in their absence pruning can be achieved by other mechanisms.
Keywords: Brain Development; Electrophysiology; Hippocampus; Microglia; Synapses.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
