

Pyrrole-based inhibitors of RND-type efflux pumps reverse antibiotic resistance and display anti-virulence potential
- PMID: 38593161
- PMCID: PMC11003683
- DOI: 10.1371/journal.ppat.1012121
Pyrrole-based inhibitors of RND-type efflux pumps reverse antibiotic resistance and display anti-virulence potential
Abstract
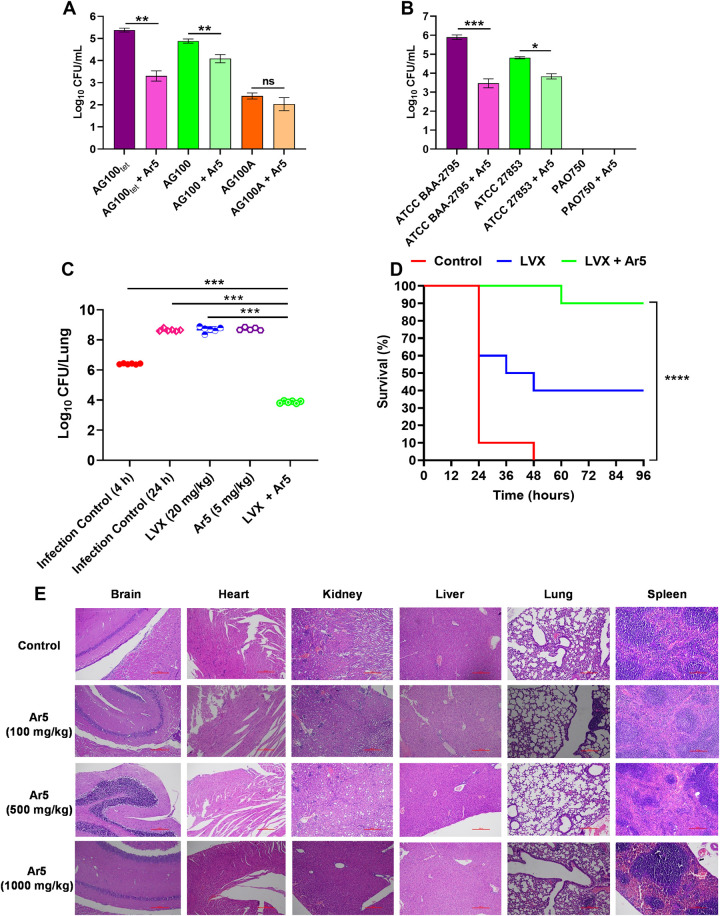
Efflux pumps of the resistance-nodulation-cell division (RND) superfamily, particularly the AcrAB-TolC, and MexAB-OprM, besides mediating intrinsic and acquired resistance, also intervene in bacterial pathogenicity. Inhibitors of such pumps could restore the activities of antibiotics and curb bacterial virulence. Here, we identify pyrrole-based compounds that boost antibiotic activity in Escherichia coli and Pseudomonas aeruginosa by inhibiting their archetype RND transporters. Molecular docking and biophysical studies revealed that the EPIs bind to AcrB. The identified efflux pump inhibitors (EPIs) inhibit the efflux of fluorescent probes, attenuate persister formation, extend post-antibiotic effect, and diminish resistant mutant development. The bacterial membranes remained intact upon exposure to the EPIs. EPIs also possess an anti-pathogenic potential and attenuate P. aeruginosa virulence in vivo. The intracellular invasion of E. coli and P. aeruginosa inside the macrophages was hampered upon treatment with the lead EPI. The excellent efficacy of the EPI-antibiotic combination was evidenced in animal lung infection and sepsis protection models. These findings indicate that EPIs discovered herein with negligible toxicity are potential antibiotic adjuvants to address life-threatening Gram-negative bacterial infections.
Copyright: © 2024 Mahey et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
N.M., R.T., R.K., R.I., N.C., S.J., and H.N. are inventors for the patent application of the EPIs disclosed in this study (Indian provisional patent Application no. 202011054425). The remaining authors declare no conflict of interest.
Figures







References
-
- Tacconelli E, Carrara E, Savoldi A, Harbarth S, Mendelson M, Monnet DL, et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. The Lancet Infectious Diseases. 2018;18(3):318–27. doi: 10.1016/S1473-3099(17)30753-3 - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
