

PRDM16 co-operates with LHX2 to shape the human brain
- PMID: 38595939
- PMCID: PMC10914218
- DOI: 10.1093/oons/kvae001
PRDM16 co-operates with LHX2 to shape the human brain
Abstract
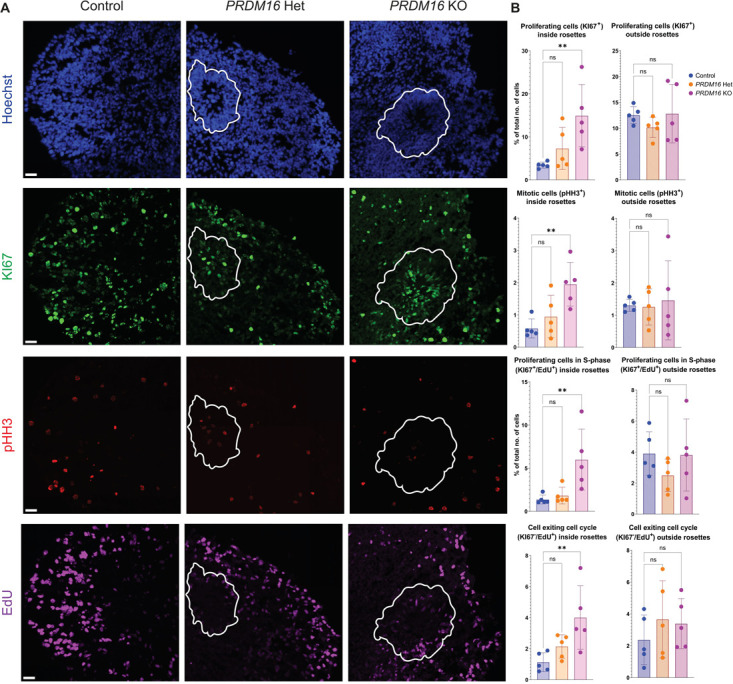
PRDM16 is a dynamic transcriptional regulator of various stem cell niches, including adipocytic, hematopoietic, cardiac progenitors, and neural stem cells. PRDM16 has been suggested to contribute to 1p36 deletion syndrome, one of the most prevalent subtelomeric microdeletion syndromes. We report a patient with a de novo nonsense mutation in the PRDM16 coding sequence, accompanied by lissencephaly and microcephaly features. Human stem cells were genetically modified to mimic this mutation, generating cortical organoids that exhibited altered cell cycle dynamics. RNA sequencing of cortical organoids at day 32 unveiled changes in cell adhesion and WNT-signaling pathways. ChIP-seq of PRDM16 identified binding sites in postmortem human fetal cortex, indicating the conservation of PRDM16 binding to developmental genes in mice and humans, potentially at enhancer sites. A shared motif between PRDM16 and LHX2 was identified and further examined through comparison with LHX2 ChIP-seq data from mice. These results suggested a collaborative partnership between PRDM16 and LHX2 in regulating a common set of genes and pathways in cortical radial glia cells, possibly via their synergistic involvement in cortical development.
Keywords: 1p36 deletion syndrome; LHX2; PRDM16; brain development; cerebral organoids; human disease model.
© The Author(s) 2024. Published by Oxford University Press. All rights reserved.
Figures





Update of
-
PRDM16 co-operates with LHX2 to shape the human brain.bioRxiv [Preprint]. 2023 Aug 12:2023.08.12.553065. doi: 10.1101/2023.08.12.553065. bioRxiv. 2023. Update in: Oxf Open Neurosci. 2024 Jan 24;3:kvae001. doi: 10.1093/oons/kvae001. PMID: 37609127 Free PMC article. Updated. Preprint.
References
-
- Chi J, Cohen P. The multifaceted roles of PRDM16: adipose biology and beyond. Trends Endocrinol Metab 2016;27:11–23 - PubMed
-
- Pinheiro I, Margueron R, Shukeir N et al. Prdm3 and Prdm16 are H3K9me1 methyltransferases required for mammalian heterochromatin integrity. Cell 2012;150:948–60 - PubMed
-
- Warner DR, Horn KH, Mudd L et al. PRDM16/MEL1: a novel Smad binding protein expressed in murine embryonic orofacial tissue. Biochim Biophys Acta 2007;1773:814–20 - PubMed
-
- Takahata M, Inoue Y, Tsuda H et al. SKI and MEL1 cooperate to inhibit transforming growth factor-beta signal in gastric cancer cells. J Biol Chem 2009;284:3334–44 - PubMed