

N-terminal signals in the SNX-BAR paralogs Vps5 and Vin1 guide endosomal coat complex formation
- PMID: 38598303
- PMCID: PMC11238075
- DOI: 10.1091/mbc.E24-01-0043
N-terminal signals in the SNX-BAR paralogs Vps5 and Vin1 guide endosomal coat complex formation
Abstract
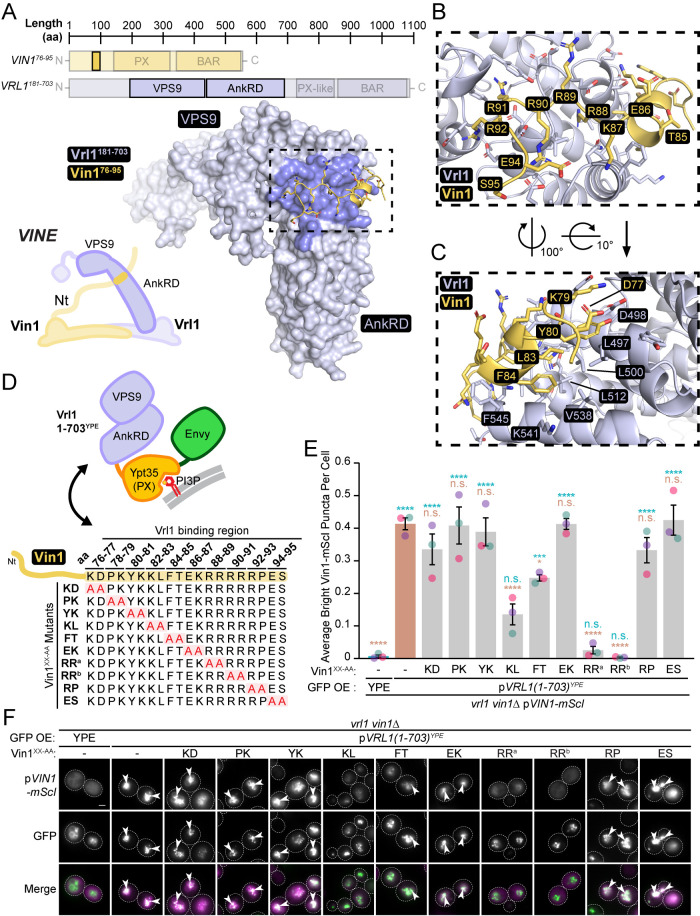
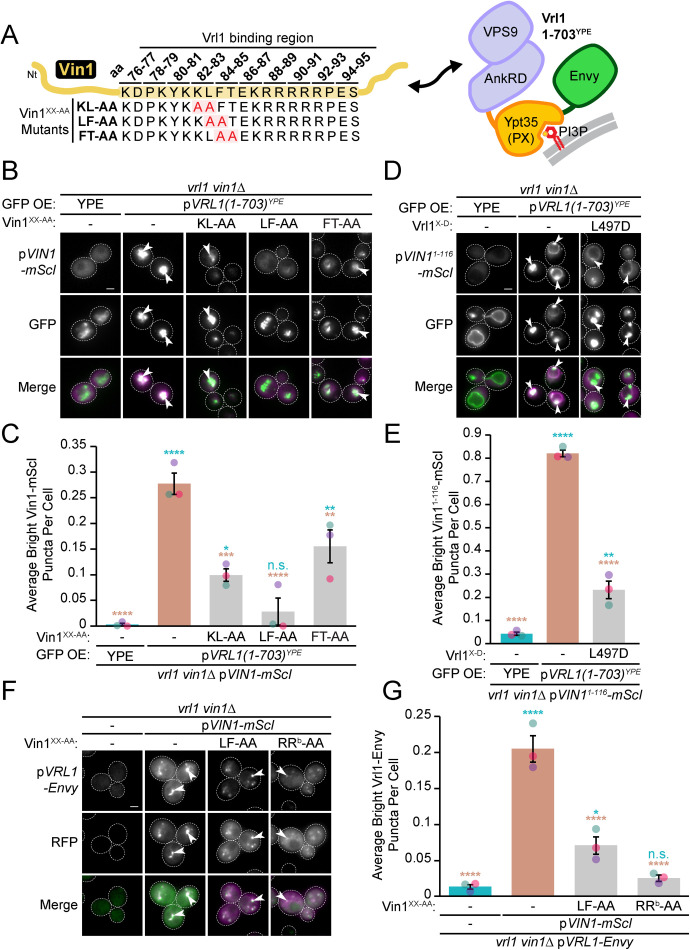
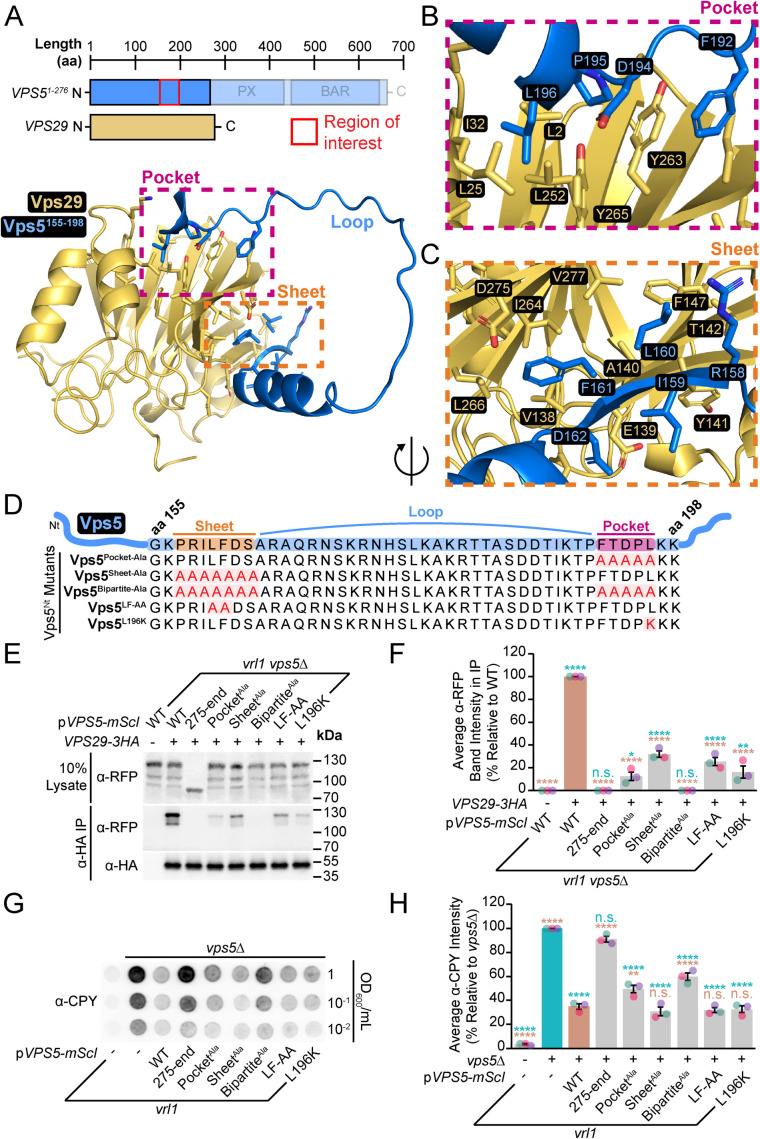
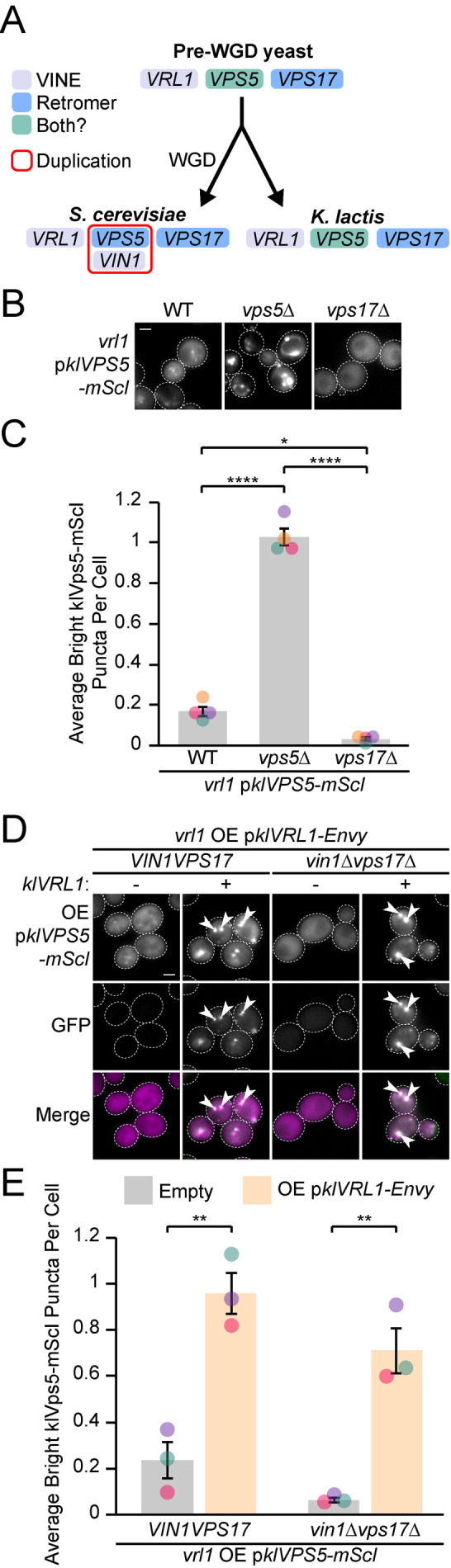
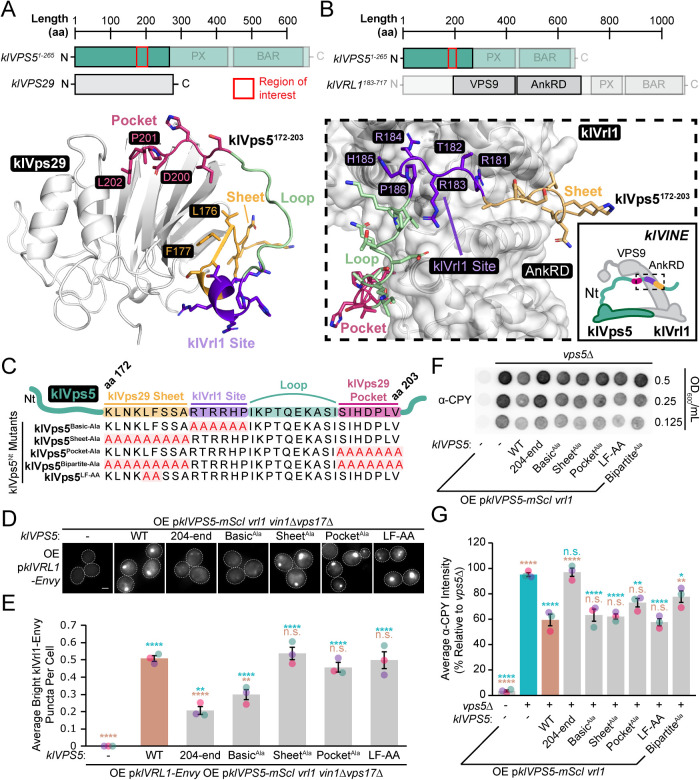
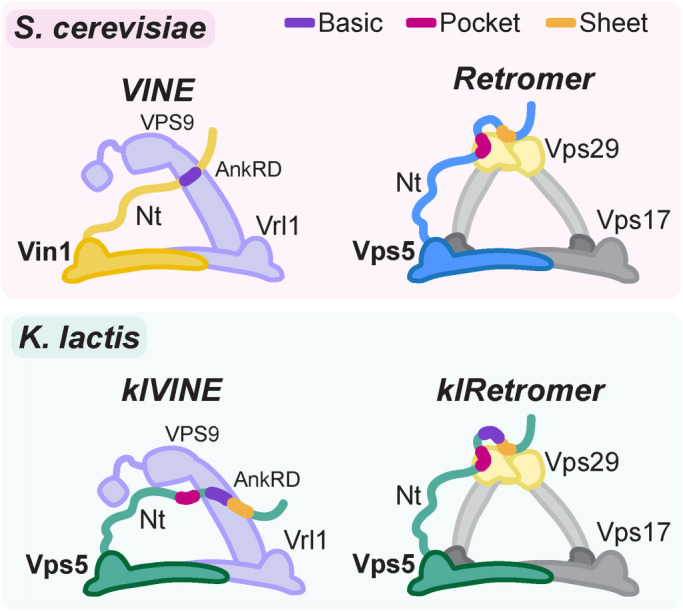
Endosomal coats incorporate membrane-binding subunits such as sorting nexin (SNX) proteins. The Saccharomyces cerevisiae SNX-BAR paralogs Vin1 and Vps5 are respective subunits of the endosomal VINE and retromer complexes whose dimerizing BAR domains are required for complex assembly and membrane association. However, a degree of promiscuity is predicted for yeast BAR-BAR pairings, and recent work has implicated the unstructured N-terminal domains of Vin1 and Vps5 in coat formation. Here, we map N-terminal signals in both SNX-BAR paralogs that contribute to the assembly and function of two distinct endosomal coats in vivo. Whereas Vin1 leverages a polybasic region and adjacent hydrophobic motif to bind Vrl1 and form VINE, the N-terminus of Vps5 interacts with the retromer subunit Vps29 at two sites, including a conserved hydrophobic pocket in Vps29 that engages other accessory proteins in humans. We also examined the sole isoform of Vps5 from the milk yeast Kluyveromyces lactis and found that ancestral yeasts may have used a nested N-terminal signal to form both VINE and retromer. Our results suggest that the specific assembly of Vps5-family SNX-BAR coats depends on inputs from unique N-terminal sequence features in addition to BAR domain coupling, expanding our understanding of endosomal coat biology.
Figures
Similar articles
-
Molecular basis for the assembly of the Vps5-Vps17 SNX-BAR proteins with Retromer.Nat Commun. 2025 Apr 15;16(1):3568. doi: 10.1038/s41467-025-58846-8. Nat Commun. 2025. PMID: 40234461 Free PMC article.
-
The VINE complex is an endosomal VPS9-domain GEF and SNX-BAR coat.Elife. 2022 Aug 8;11:e77035. doi: 10.7554/eLife.77035. Elife. 2022. PMID: 35938928 Free PMC article.
-
Retromer oligomerization drives SNX-BAR coat assembly and membrane constriction.EMBO J. 2023 Jan 16;42(2):e112287. doi: 10.15252/embj.2022112287. EMBO J. 2023. PMID: 36644906 Free PMC article.
-
You can go your own way: SNX-BAR coat complexes direct traffic at late endosomes.Curr Opin Cell Biol. 2022 Jun;76:102087. doi: 10.1016/j.ceb.2022.102087. Epub 2022 May 12. Curr Opin Cell Biol. 2022. PMID: 35569261 Review.
-
Updated Insight into the Physiological and Pathological Roles of the Retromer Complex.Int J Mol Sci. 2017 Jul 25;18(8):1601. doi: 10.3390/ijms18081601. Int J Mol Sci. 2017. PMID: 28757549 Free PMC article. Review.
Cited by
-
Molecular basis for the assembly of the Vps5-Vps17 SNX-BAR proteins with Retromer.Nat Commun. 2025 Apr 15;16(1):3568. doi: 10.1038/s41467-025-58846-8. Nat Commun. 2025. PMID: 40234461 Free PMC article.
-
Identification of a VPS29 isoform with restricted association to Retriever and Retromer accessory proteins through autoinhibition.Proc Natl Acad Sci U S A. 2025 Jul 8;122(27):e2501111122. doi: 10.1073/pnas.2501111122. Epub 2025 Jun 30. Proc Natl Acad Sci U S A. 2025. PMID: 40587794 Free PMC article.
References
-
- Bean BDM, Davey M, Conibear E (2017). Cargo selectivity of yeast sorting nexins. Traffic 18, 110–122. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
