

Emergence of fractal geometries in the evolution of a metabolic enzyme
- PMID: 38600380
- PMCID: PMC11041685
- DOI: 10.1038/s41586-024-07287-2
Emergence of fractal geometries in the evolution of a metabolic enzyme
Abstract
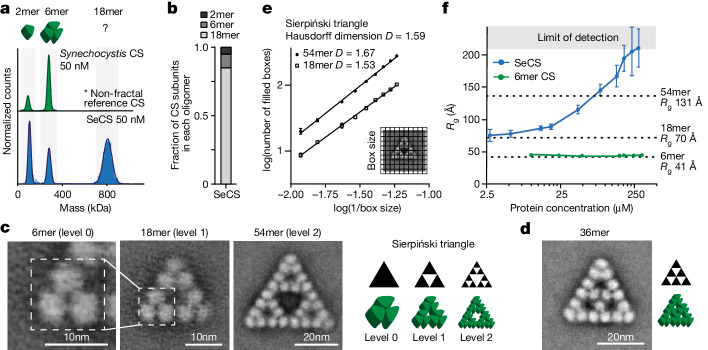
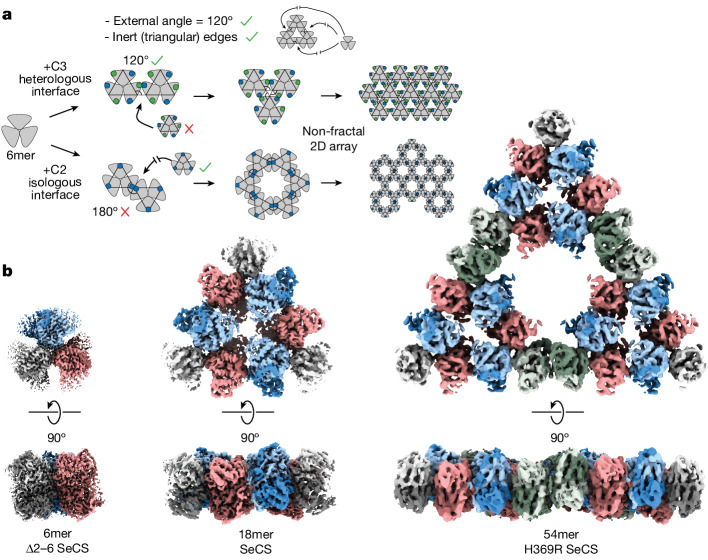
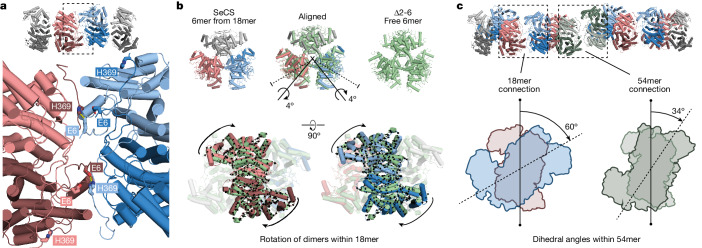
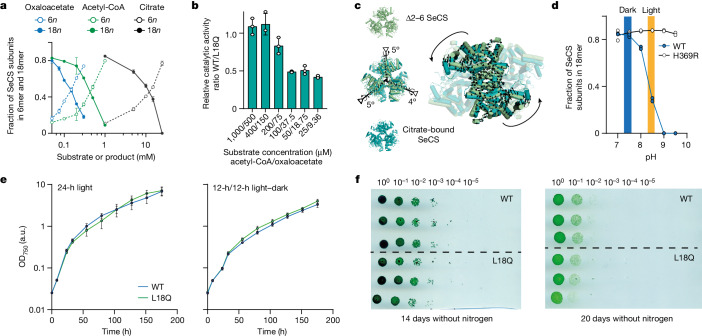
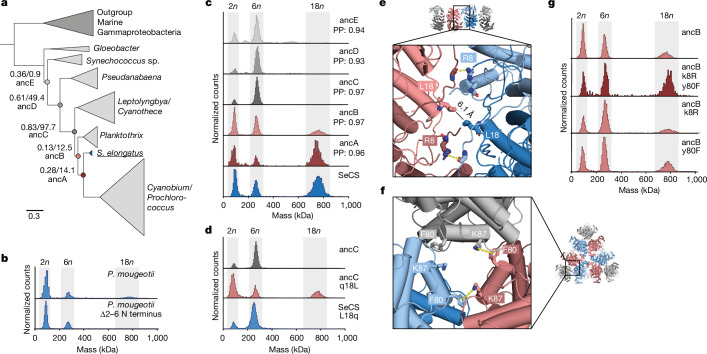
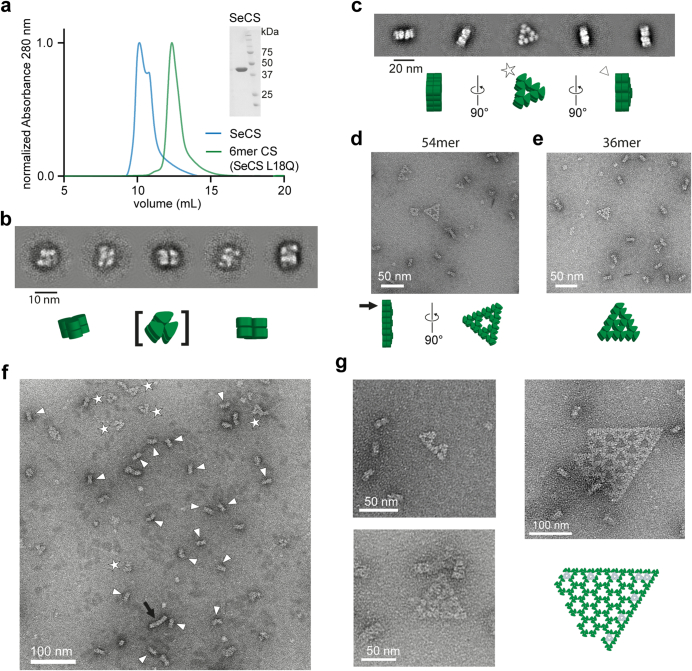
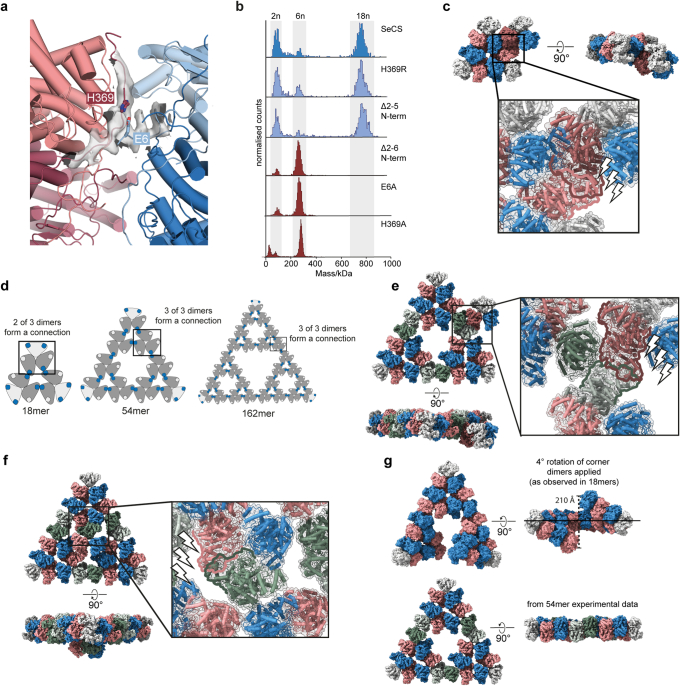
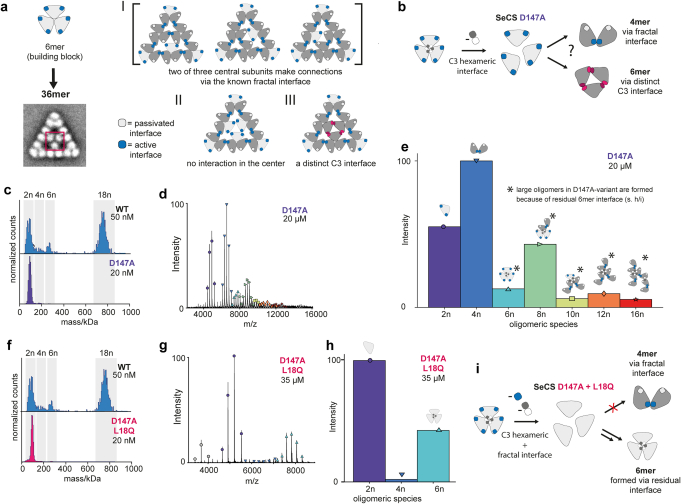
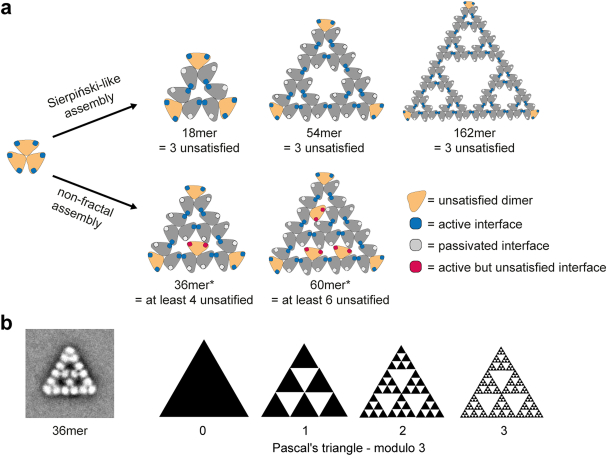
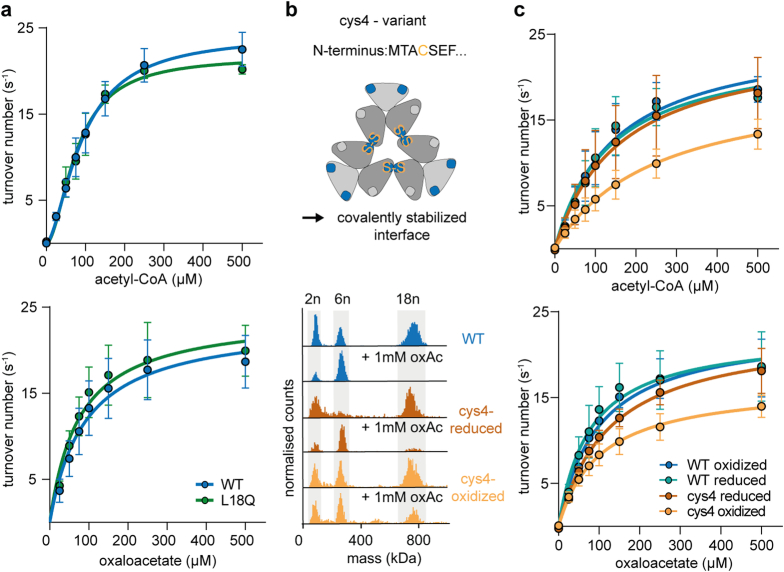
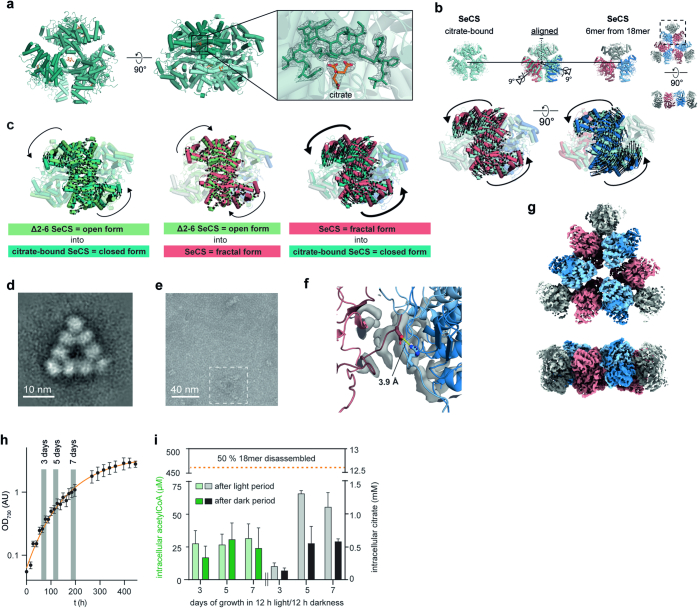
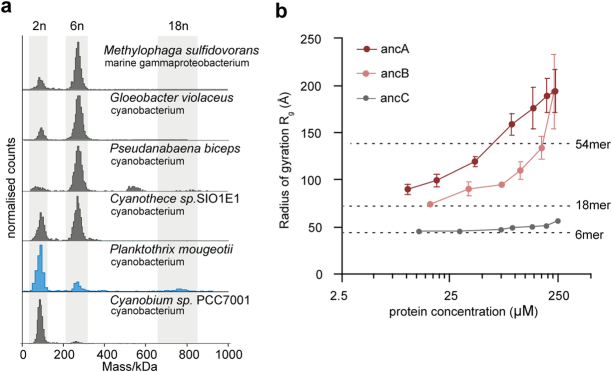
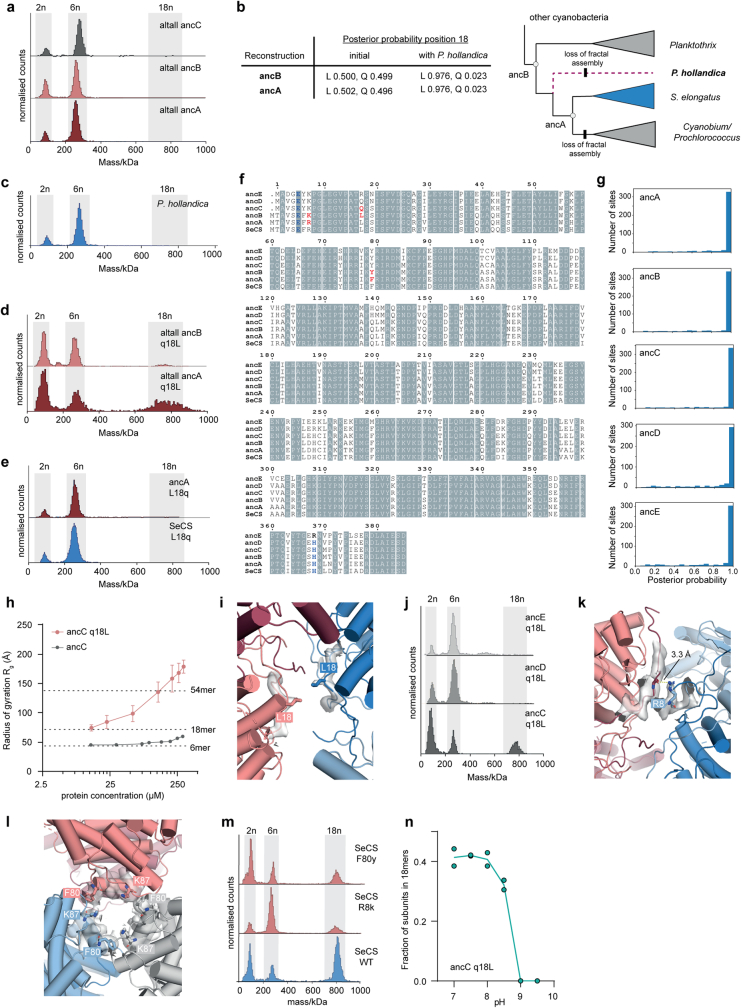
Fractals are patterns that are self-similar across multiple length-scales1. Macroscopic fractals are common in nature2-4; however, so far, molecular assembly into fractals is restricted to synthetic systems5-12. Here we report the discovery of a natural protein, citrate synthase from the cyanobacterium Synechococcus elongatus, which self-assembles into Sierpiński triangles. Using cryo-electron microscopy, we reveal how the fractal assembles from a hexameric building block. Although different stimuli modulate the formation of fractal complexes and these complexes can regulate the enzymatic activity of citrate synthase in vitro, the fractal may not serve a physiological function in vivo. We use ancestral sequence reconstruction to retrace how the citrate synthase fractal evolved from non-fractal precursors, and the results suggest it may have emerged as a harmless evolutionary accident. Our findings expand the space of possible protein complexes and demonstrate that intricate and regulatable assemblies can evolve in a single substitution.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Mandelbrot, B. B. The Fractal Geometry of Nature. Vol. 1 (W. H. Freeman and Co., 1982).
-
- Du, J. X., Zhai, C. M. & Wang, Q. P. Recognition of plant leaf image based on fractal dimension features. Neurocomputing116, 150–156 (2013).10.1016/j.neucom.2012.03.028 - DOI
-
- Tarboton, D. G., Bras, R. L. & Rodriguez-Iturbe, I. The fractal nature of river networks. Water Resour. Res.24, 1317–1322 (1988).10.1029/WR024i008p01317 - DOI
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
