

Human gene regulatory evolution is driven by the divergence of regulatory element function in both cis and trans
- PMID: 38604126
- PMCID: PMC11019363
- DOI: 10.1016/j.xgen.2024.100536
Human gene regulatory evolution is driven by the divergence of regulatory element function in both cis and trans
Abstract
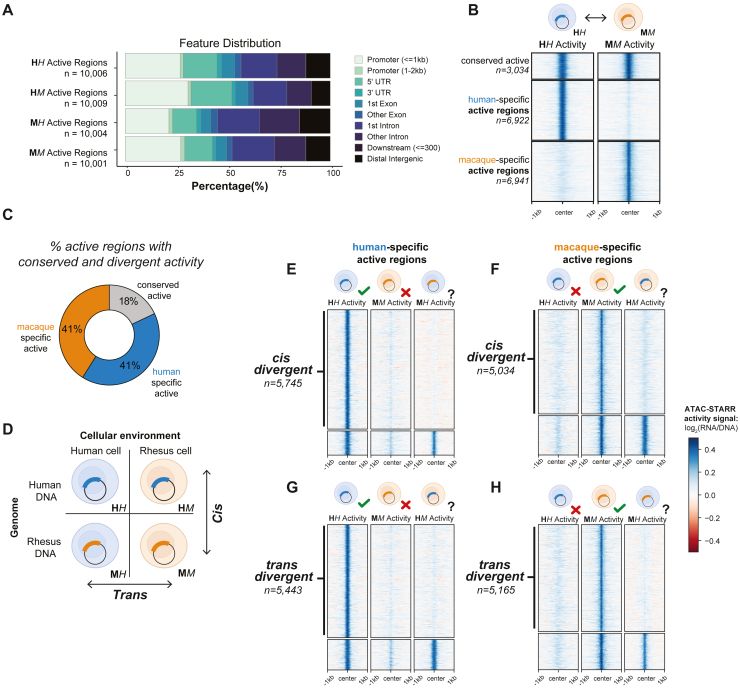
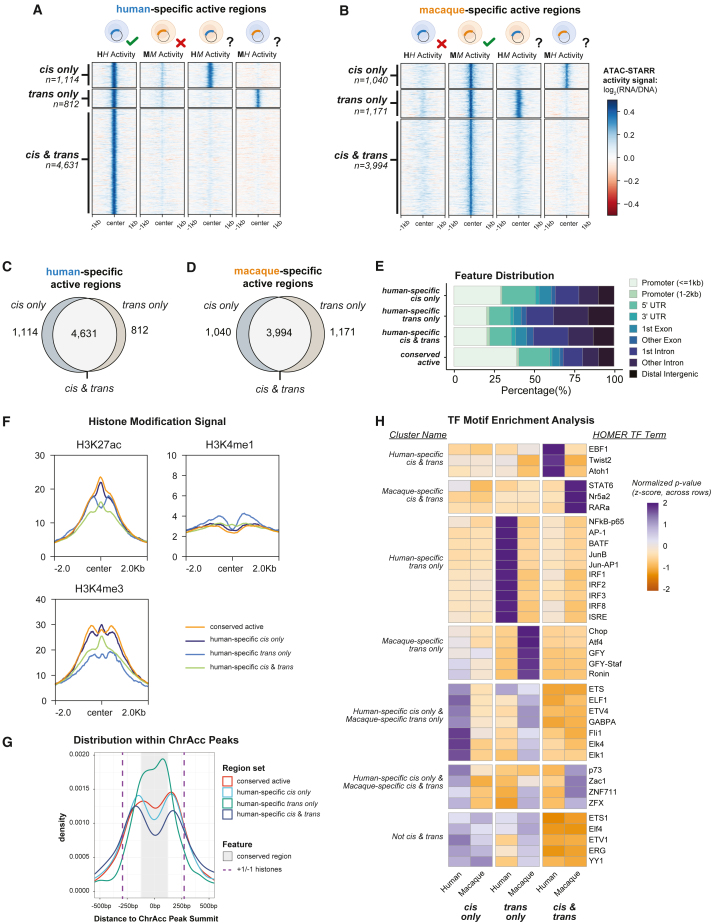
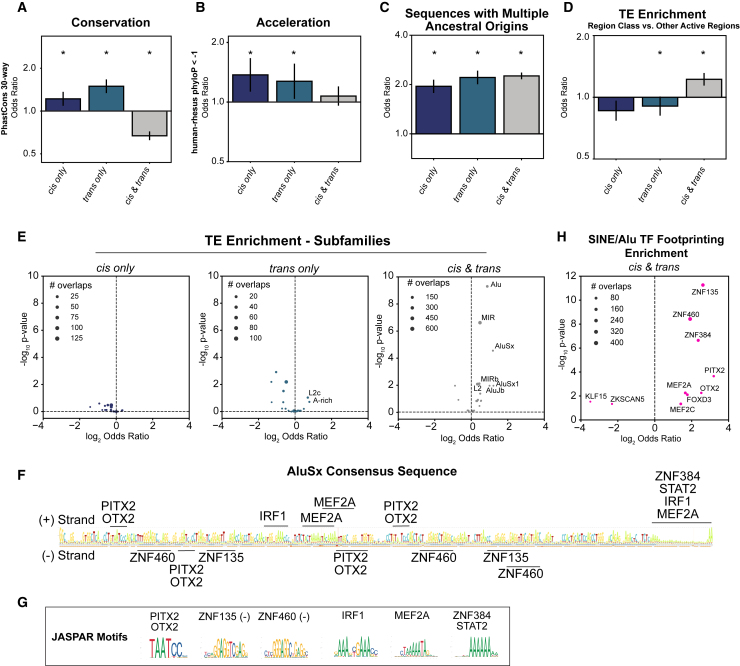
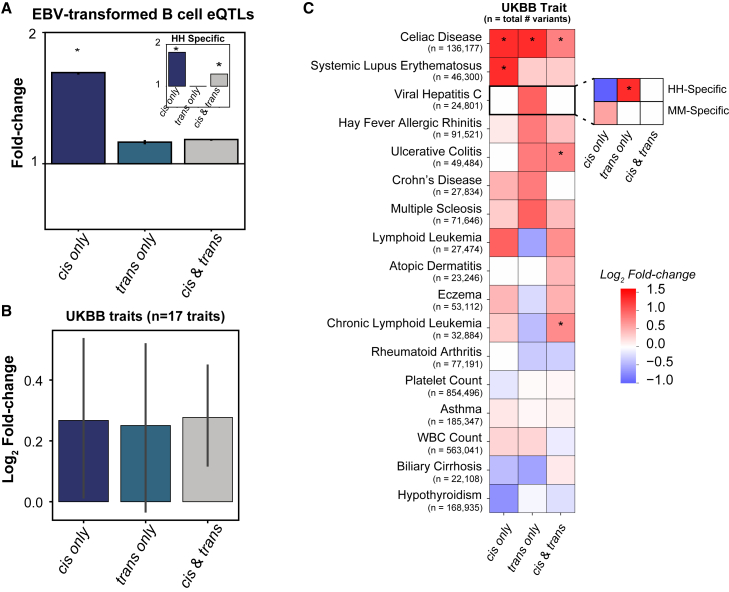
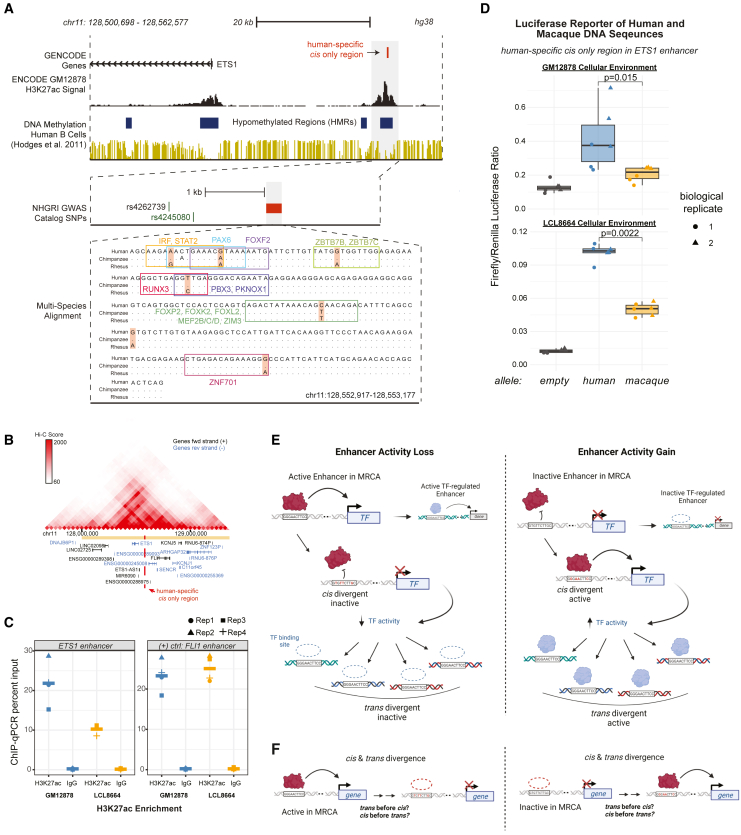
Gene regulatory divergence between species can result from cis-acting local changes to regulatory element DNA sequences or global trans-acting changes to the regulatory environment. Understanding how these mechanisms drive regulatory evolution has been limited by challenges in identifying trans-acting changes. We present a comprehensive approach to directly identify cis- and trans-divergent regulatory elements between human and rhesus macaque lymphoblastoid cells using assay for transposase-accessible chromatin coupled to self-transcribing active regulatory region (ATAC-STARR) sequencing. In addition to thousands of cis changes, we discover an unexpected number (∼10,000) of trans changes and show that cis and trans elements exhibit distinct patterns of sequence divergence and function. We further identify differentially expressed transcription factors that underlie ∼37% of trans differences and trace how cis changes can produce cascades of trans changes. Overall, we find that most divergent elements (67%) experienced changes in both cis and trans, revealing a substantial role for trans divergence-alone and together with cis changes-in regulatory differences between species.
Keywords: DNA regulatory elements; chromatin accessibility; comparative genomics; enhancer activity; functional genomics; gene regulation; human evolution; lymphoblastoid cell lines; massively parallel reporter assays; transcription factors.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures



Update of
-
Human gene regulatory evolution is driven by the divergence of regulatory element function in both cis and trans.bioRxiv [Preprint]. 2023 Feb 15:2023.02.14.528376. doi: 10.1101/2023.02.14.528376. bioRxiv. 2023. Update in: Cell Genom. 2024 Apr 10;4(4):100536. doi: 10.1016/j.xgen.2024.100536. PMID: 36824965 Free PMC article. Updated. Preprint.
Similar articles
-
Transforming our understanding of species-specific gene regulation.Cell Genom. 2024 Apr 10;4(4):100540. doi: 10.1016/j.xgen.2024.100540. Cell Genom. 2024. PMID: 38604125 Free PMC article.
-
Human gene regulatory evolution is driven by the divergence of regulatory element function in both cis and trans.bioRxiv [Preprint]. 2023 Feb 15:2023.02.14.528376. doi: 10.1101/2023.02.14.528376. bioRxiv. 2023. Update in: Cell Genom. 2024 Apr 10;4(4):100536. doi: 10.1016/j.xgen.2024.100536. PMID: 36824965 Free PMC article. Updated. Preprint.
-
ATAC-STARR-seq reveals transcription factor-bound activators and silencers within chromatin-accessible regions of the human genome.Genome Res. 2022 Aug 25;32(8):1529-1541. doi: 10.1101/gr.276766.122. Genome Res. 2022. PMID: 35858748 Free PMC article.
-
Understanding how cis-regulatory function is encoded in DNA sequence using massively parallel reporter assays and designed sequences.Genomics. 2015 Sep;106(3):165-170. doi: 10.1016/j.ygeno.2015.06.003. Epub 2015 Jun 10. Genomics. 2015. PMID: 26072432 Free PMC article. Review.
-
Cis-regulatory elements: molecular mechanisms and evolutionary processes underlying divergence.Nat Rev Genet. 2011 Dec 6;13(1):59-69. doi: 10.1038/nrg3095. Nat Rev Genet. 2011. PMID: 22143240 Review.
Cited by
-
Addressing missing context in regulatory variation across primate evolution.ArXiv [Preprint]. 2025 Apr 2:arXiv:2504.02081v1. ArXiv. 2025. Update in: Curr Opin Genet Dev. 2025 Aug;93:102374. doi: 10.1016/j.gde.2025.102374. PMID: 40236837 Free PMC article. Updated. Preprint.
-
Transforming our understanding of species-specific gene regulation.Cell Genom. 2024 Apr 10;4(4):100540. doi: 10.1016/j.xgen.2024.100540. Cell Genom. 2024. PMID: 38604125 Free PMC article.
-
Novelty versus innovation of gene regulatory elements in human evolution and disease.Curr Opin Genet Dev. 2025 Feb;90:102279. doi: 10.1016/j.gde.2024.102279. Epub 2024 Nov 26. Curr Opin Genet Dev. 2025. PMID: 39591813 Review.
-
Uncovering methylation-dependent genetic effects on regulatory element function in diverse genomes.Genome Res. 2025 Aug 1;35(8):1781-1793. doi: 10.1101/gr.279957.124. Genome Res. 2025. PMID: 40659498 Free PMC article.
-
Transcription of Endogenous Retroviruses: Broad and Precise Mechanisms of Control.Viruses. 2024 Aug 17;16(8):1312. doi: 10.3390/v16081312. Viruses. 2024. PMID: 39205286 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
