

Representing stimulus motion with waves in adaptive neural fields
- PMID: 38607466
- PMCID: PMC11802407
- DOI: 10.1007/s10827-024-00869-z
Representing stimulus motion with waves in adaptive neural fields
Abstract
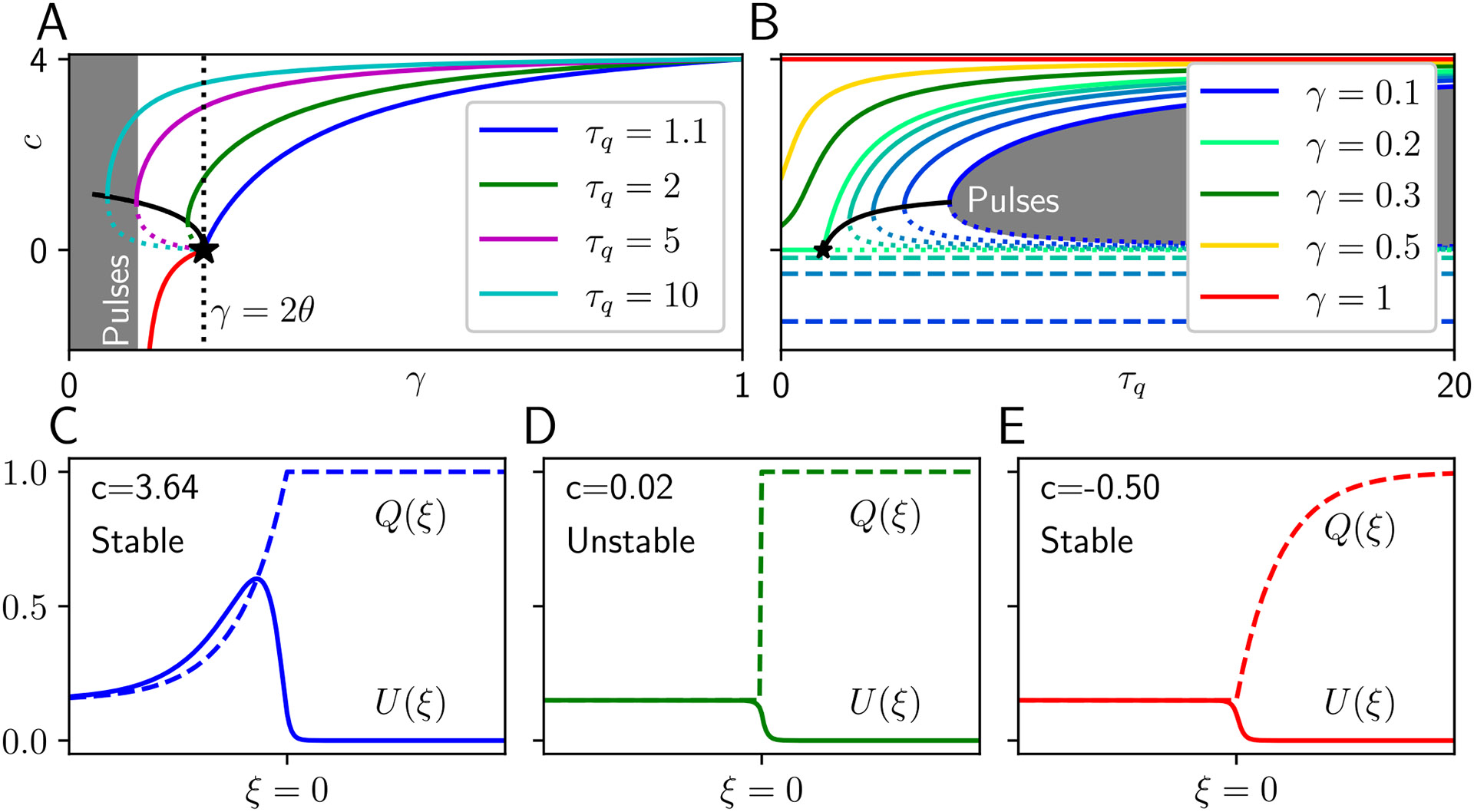
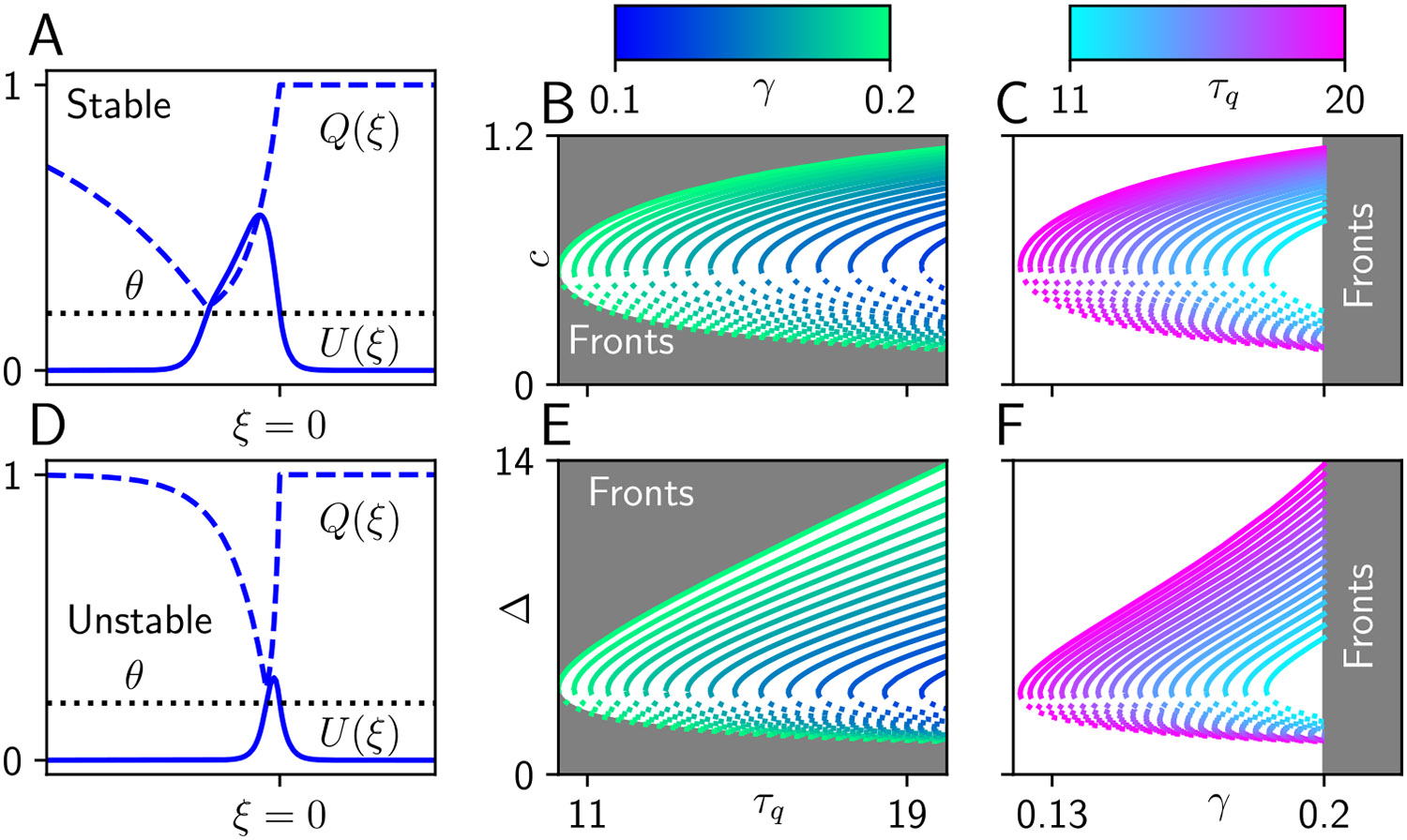
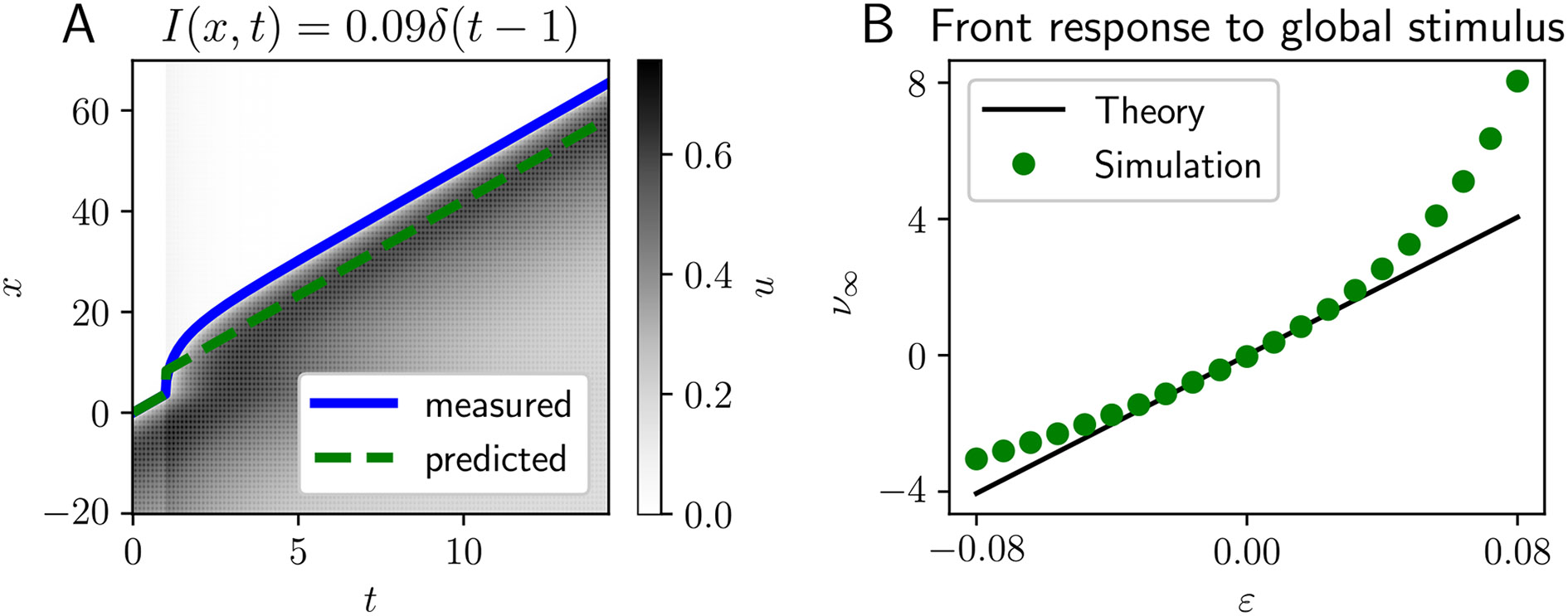
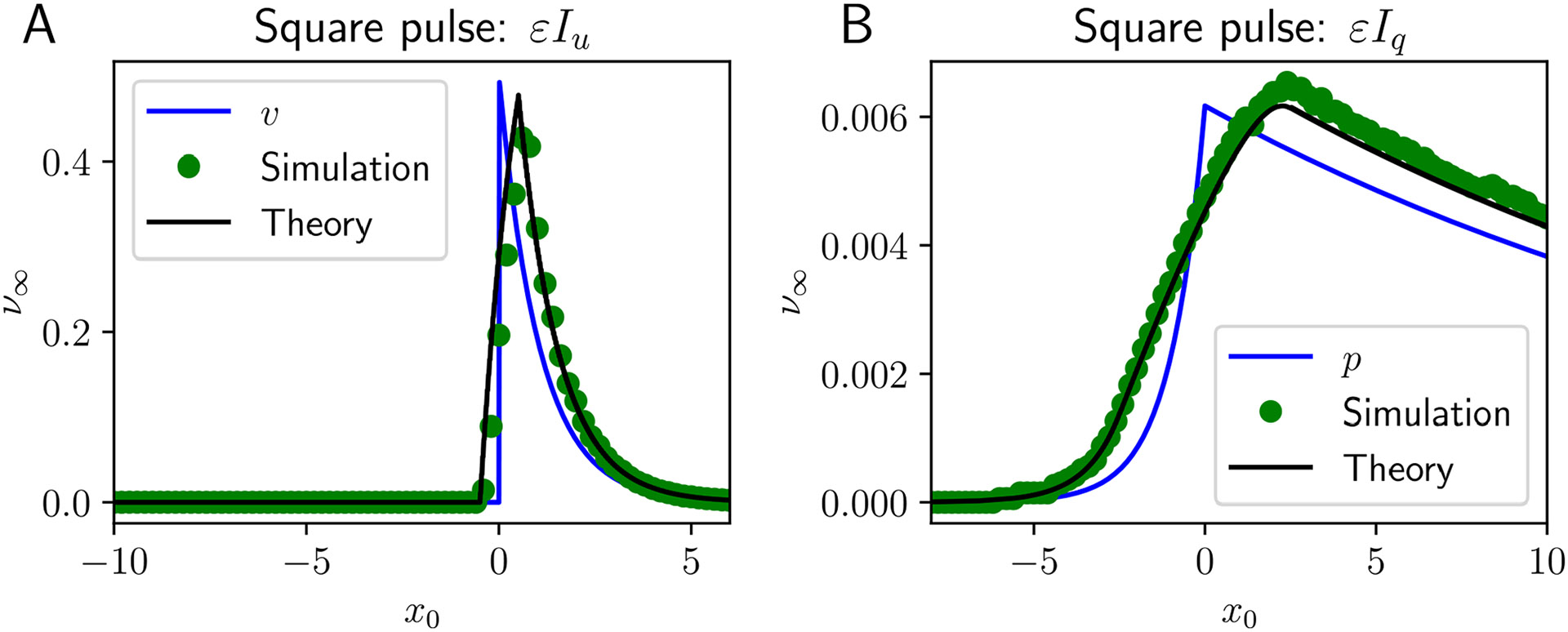
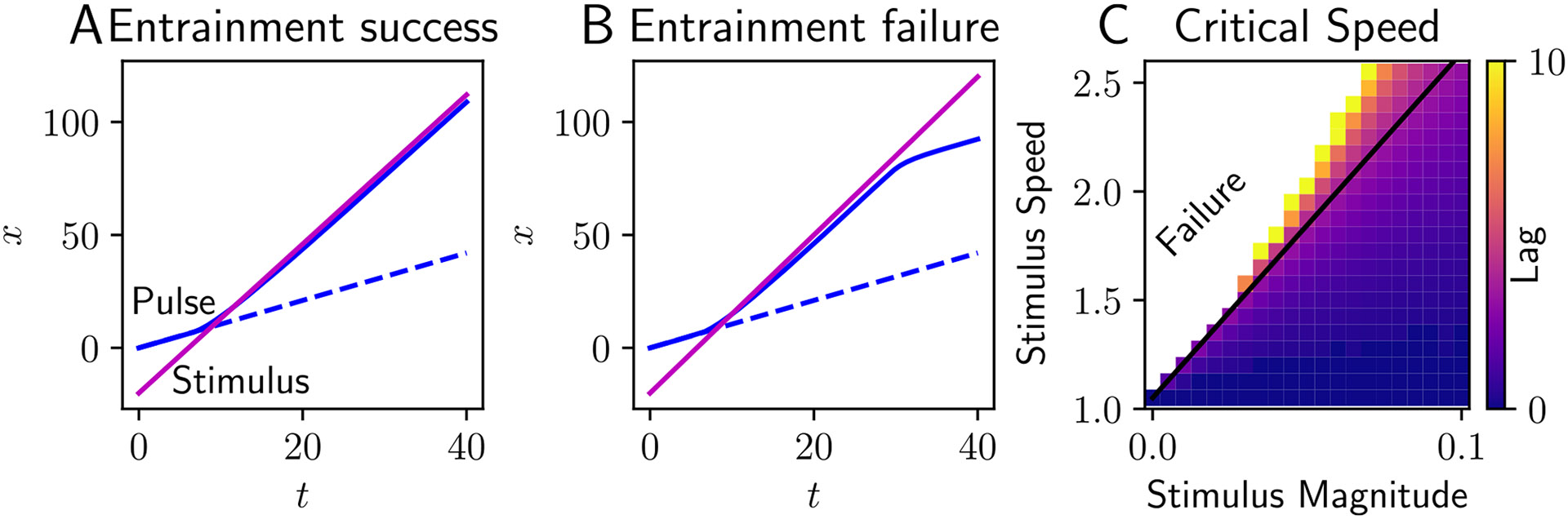
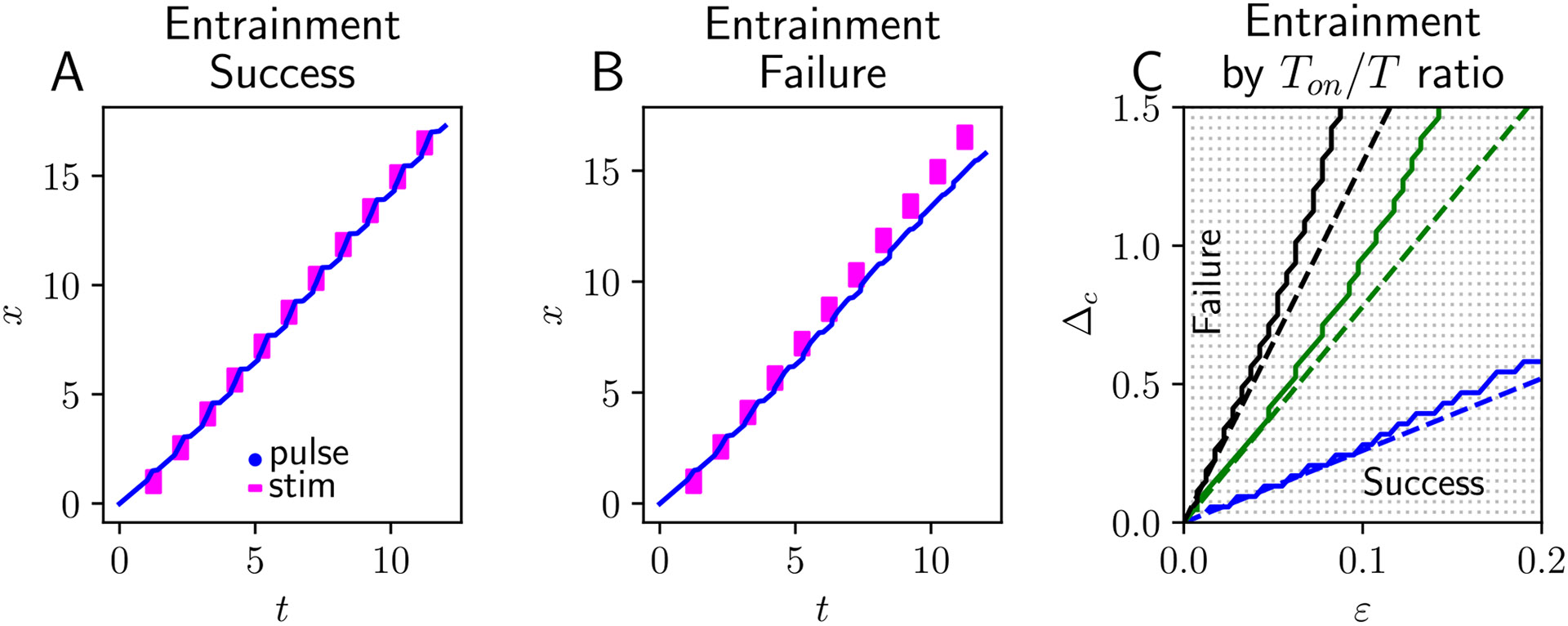
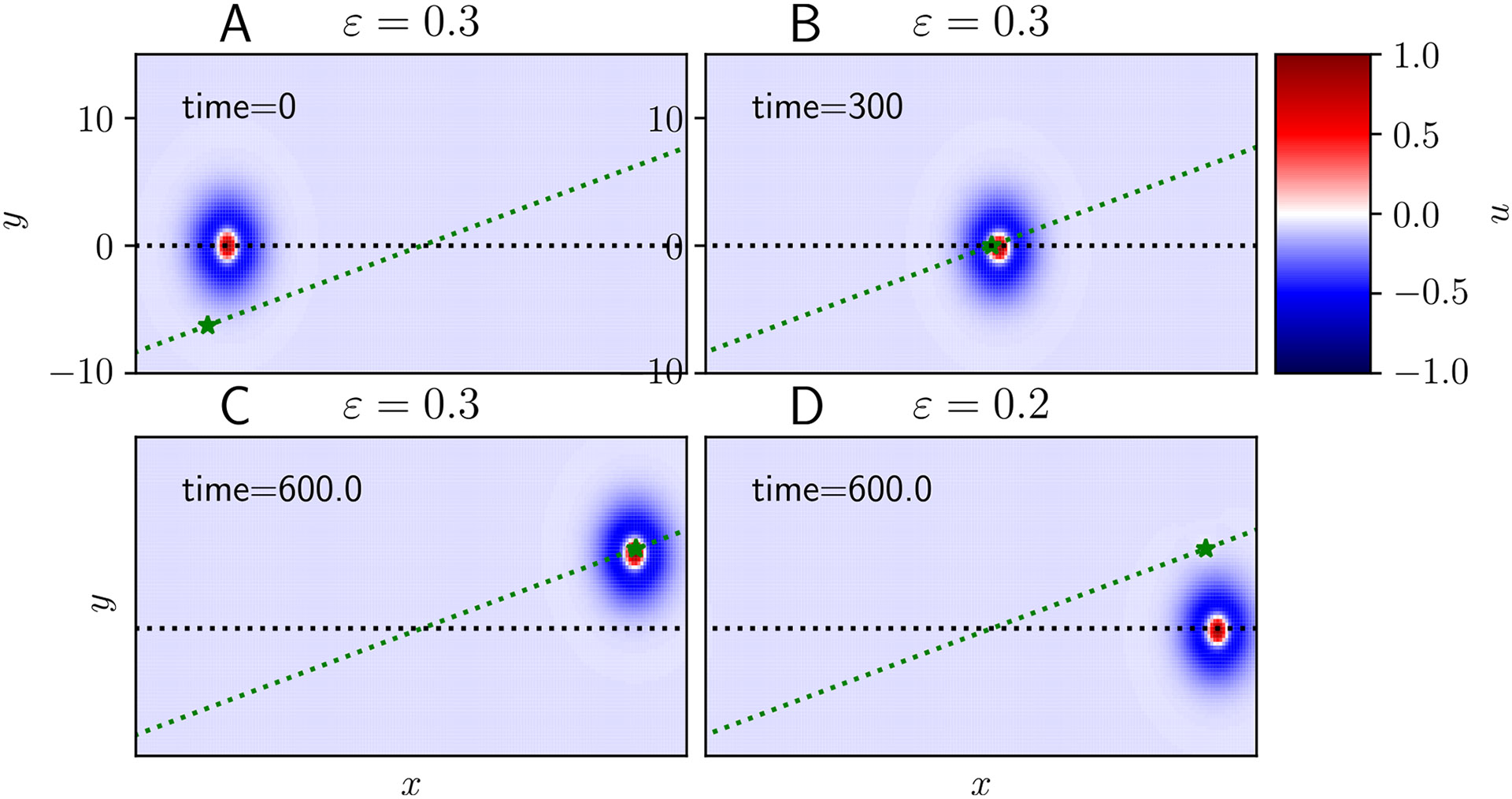
Traveling waves of neural activity emerge in cortical networks both spontaneously and in response to stimuli. The spatiotemporal structure of waves can indicate the information they encode and the physiological processes that sustain them. Here, we investigate the stimulus-response relationships of traveling waves emerging in adaptive neural fields as a model of visual motion processing. Neural field equations model the activity of cortical tissue as a continuum excitable medium, and adaptive processes provide negative feedback, generating localized activity patterns. Synaptic connectivity in our model is described by an integral kernel that weakens dynamically due to activity-dependent synaptic depression, leading to marginally stable traveling fronts (with attenuated backs) or pulses of a fixed speed. Our analysis quantifies how weak stimuli shift the relative position of these waves over time, characterized by a wave response function we obtain perturbatively. Persistent and continuously visible stimuli model moving visual objects. Intermittent flashes that hop across visual space can produce the experience of smooth apparent visual motion. Entrainment of waves to both kinds of moving stimuli are well characterized by our theory and numerical simulations, providing a mechanistic description of the perception of visual motion.
Keywords: Neural field; Synaptic depression; Traveling waves; Visual object motion.
© 2024. The Author(s), under exclusive licence to Springer Science+Business Media, LLC, part of Springer Nature.
Conflict of interest statement
Figures
Update of
-
Representing stimulus motion with waves in adaptive neural fields.ArXiv [Preprint]. 2023 Dec 11:arXiv:2312.06100v1. ArXiv. 2023. Update in: J Comput Neurosci. 2024 May;52(2):145-164. doi: 10.1007/s10827-024-00869-z. PMID: 38168459 Free PMC article. Updated. Preprint.
Similar articles
-
Representing stimulus motion with waves in adaptive neural fields.ArXiv [Preprint]. 2023 Dec 11:arXiv:2312.06100v1. ArXiv. 2023. Update in: J Comput Neurosci. 2024 May;52(2):145-164. doi: 10.1007/s10827-024-00869-z. PMID: 38168459 Free PMC article. Updated. Preprint.
-
Suppressive Traveling Waves Shape Representations of Illusory Motion in Primary Visual Cortex of Awake Primate.J Neurosci. 2019 May 29;39(22):4282-4298. doi: 10.1523/JNEUROSCI.2792-18.2019. Epub 2019 Mar 18. J Neurosci. 2019. PMID: 30886010 Free PMC article.
-
Stimulus motion propels traveling waves in binocular rivalry.PLoS One. 2007 Aug 15;2(8):e739. doi: 10.1371/journal.pone.0000739. PLoS One. 2007. PMID: 17710139 Free PMC article.
-
Traveling waves and the processing of weakly tuned inputs in a cortical network module.J Comput Neurosci. 1997 Jan;4(1):57-77. doi: 10.1023/a:1008816611284. J Comput Neurosci. 1997. PMID: 9046452
-
Traveling waves in visual cortex.Neuron. 2012 Jul 26;75(2):218-29. doi: 10.1016/j.neuron.2012.06.029. Neuron. 2012. PMID: 22841308 Review.
References
-
- Abouzeid A, & Ermentrout B (2009). Type-ii phase resetting curve is optimal for stochastic synchrony. Physical Review E, 80(1), 011911. - PubMed
-
- Alamia A, & VanRullen R (2023). A traveling waves perspective on temporal binding. Journal of Cognitive Neuroscience, pp 1–9. - PubMed
-
- Amari S. (1977). Dynamics of pattern formation in lateral-inhibition type neural fields. Biological Cybernetics, 27(2), 77–87. - PubMed
-
- Anstis SM (1980). The perception of apparent movement. Philosophical Transactions of the Royal Society of London B, Biological Sciences, 290(1038), 153–168. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
