

An electrostatic cluster guides Aβ40 fibril formation in sporadic and Dutch-type cerebral amyloid angiopathy
- PMID: 38615725
- PMCID: PMC11162928
- DOI: 10.1016/j.jsb.2024.108092
An electrostatic cluster guides Aβ40 fibril formation in sporadic and Dutch-type cerebral amyloid angiopathy
Abstract
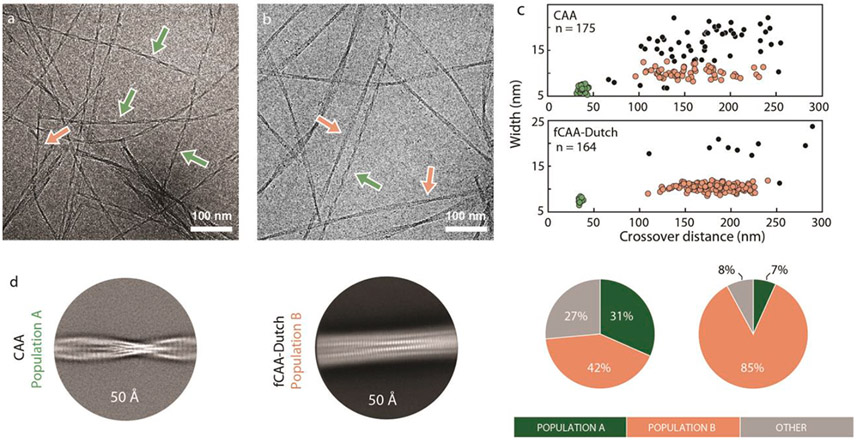
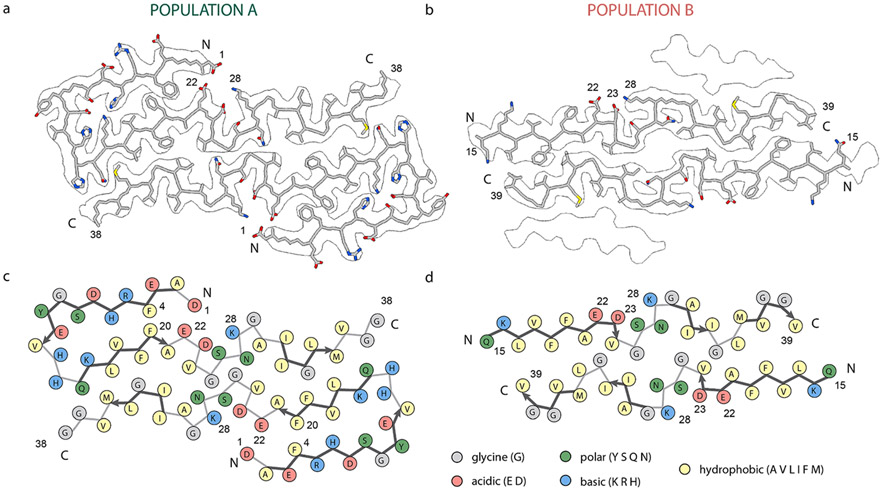
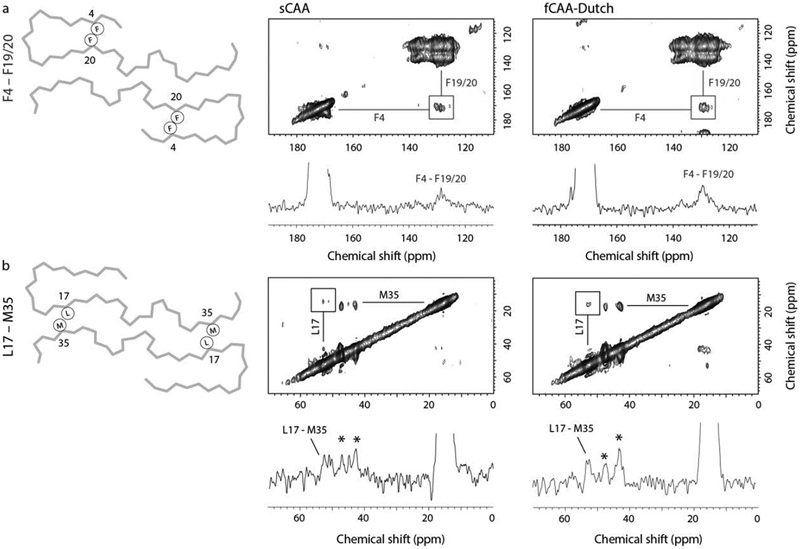
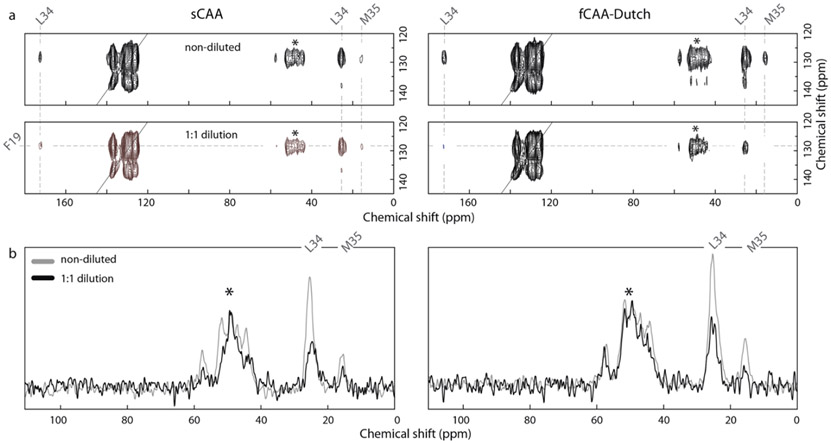
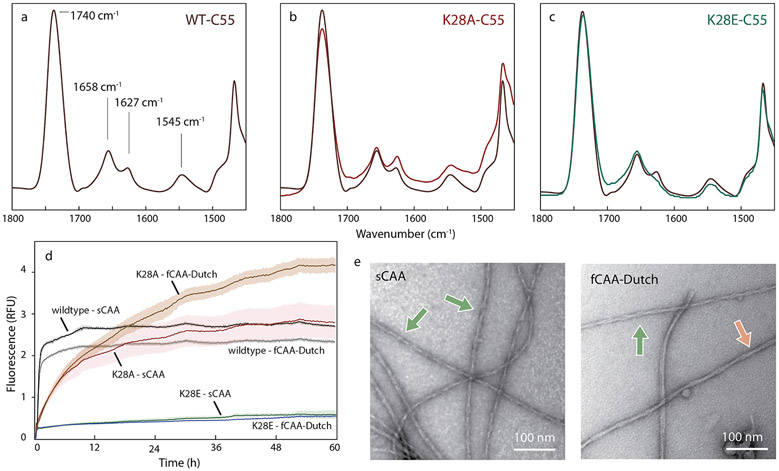
Cerebral amyloid angiopathy (CAA) is associated with the accumulation of fibrillar Aβ peptides upon and within the cerebral vasculature, which leads to loss of vascular integrity and contributes to disease progression in Alzheimer's disease (AD). We investigate the structure of human-derived Aβ40 fibrils obtained from patients diagnosed with sporadic or familial Dutch-type (E22Q) CAA. Using cryo-EM, two primary structures are identified containing elements that have not been observed in in vitro Aβ40 fibril structures. One population has an ordered N-terminal fold comprised of two β-strands stabilized by electrostatic interactions involving D1, E22, D23 and K28. This charged cluster is disrupted in the second population, which exhibits a disordered N-terminus and is favored in fibrils derived from the familial Dutch-type CAA patient. These results illustrate differences between human-derived CAA and AD fibrils, and how familial CAA mutations can guide fibril formation.
Keywords: Alzheimer's disease; Amyloid-β peptide; Cerebral amyloid angiopathy; Cryo-EM; Solid-state NMR; Vascular fibrils.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: William E. Van Nostrand, Steven O. Smith reports financial support was provided by National Institutes of Health. Co-author S.C. serves on the editoral board of the Journal of Structural Biology If there are other authors, they declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures

Similar articles
-
Aβ40 Fibril Assembly on Human Cerebral Smooth Muscle Cells Impairs Cell Viability.Biochemistry. 2025 Mar 4;64(5):1065-1078. doi: 10.1021/acs.biochem.4c00478. Epub 2025 Jan 6. Biochemistry. 2025. PMID: 39763333
-
Insights into Cerebral Amyloid Angiopathy Type 1 and Type 2 from Comparisons of the Fibrillar Assembly and Stability of the Aβ40-Iowa and Aβ40-Dutch Peptides.Biochemistry. 2022 Jun 21;61(12):1181-1198. doi: 10.1021/acs.biochem.1c00781. Epub 2022 Jun 6. Biochemistry. 2022. PMID: 35666749 Free PMC article.
-
Anti-Parallel β-Hairpin Structure in Soluble Aβ Oligomers of Aβ40-Dutch and Aβ40-Iowa.Int J Mol Sci. 2021 Jan 27;22(3):1225. doi: 10.3390/ijms22031225. Int J Mol Sci. 2021. PMID: 33513738 Free PMC article.
-
Hereditary and sporadic forms of abeta-cerebrovascular amyloidosis and relevant transgenic mouse models.Int J Mol Sci. 2009 Apr 23;10(4):1872-1895. doi: 10.3390/ijms10041872. Int J Mol Sci. 2009. PMID: 19468344 Free PMC article. Review.
-
Cerebral amyloid angiopathy.Prog Mol Biol Transl Sci. 2012;107:41-78. doi: 10.1016/B978-0-12-385883-2.00006-0. Prog Mol Biol Transl Sci. 2012. PMID: 22482447 Review.
Cited by
-
Cerebral Proteomic Changes in the rTg-D Rat Model of Cerebral Amyloid Angiopathy Type-2 With Cortical Microhemorrhages and Cognitive Impairments.Neurosci Insights. 2024 Oct 8;19:26331055241288172. doi: 10.1177/26331055241288172. eCollection 2024. Neurosci Insights. 2024. PMID: 39386146 Free PMC article.
-
Membrane-assisted Aβ40 aggregation pathways.Cell Rep Phys Sci. 2025 Feb 19;6(2):102436. doi: 10.1016/j.xcrp.2025.102436. Epub 2025 Feb 11. Cell Rep Phys Sci. 2025. PMID: 40083905 Free PMC article.
-
Cryo-EM evidence for a common factor in Alzheimer's and other neurodegenerations.bioRxiv [Preprint]. 2025 May 15:2025.05.13.653829. doi: 10.1101/2025.05.13.653829. bioRxiv. 2025. PMID: 40463114 Free PMC article. Preprint.
-
Medin drives Aβ40 to adopt Aβ42-like fibril polymorphs in vitro.bioRxiv [Preprint]. 2025 Jul 20:2025.07.17.665283. doi: 10.1101/2025.07.17.665283. bioRxiv. 2025. PMID: 40791487 Free PMC article. Preprint.
-
Ligands for Protein Fibrils of Amyloid-β, α-Synuclein, and Tau.Chem Rev. 2025 Jun 11;125(11):5282-5348. doi: 10.1021/acs.chemrev.4c00838. Epub 2025 May 6. Chem Rev. 2025. PMID: 40327808 Free PMC article. Review.
References
-
- Alvarez X, Miguel-Hidalgo J, Fernandez-Novoa L, Cacabelos R, 1997. Intrahippocampal injections of the beta-amyloid 1–28 fragment induces behavioral deficits in rats. Methods Find. Exp. Clin. Pharmacol 19, 471–479. - PubMed
-
- Bakrania P, Hall G, Bouter Y, Bouter C, Beindorff N, Cowan R, Davies S, Price J, Mpamhanga C, Love E, Matthews D, Carr MD, Bayer TA, 2022. Discovery of a novel pseudo β-hairpin structure of N-truncated amyloid-β for use as a vaccine against Alzheimer’s disease. Mol. Psychiatry 27, 840–848. 10.1038/S41380-021-01385-7. - DOI - PubMed
-
- Bohrmann B, Baumann K, Benz J, Gerber F, Huber W, Knoflach F, Messer J, Oroszlan K, Rauchenberger R, Richter WF, Rothe C, Urban M, Bardroff M, Winter M, Nordstedt C, Loetscher H, 2012. Gantenerumab: a novel human anti-Aβ antibody demonstrates sustained cerebral amyloid-β binding and elicits cell-mediated removal of human amyloid-β. J. Alzheimer’s Disease 28, 49–69. 10.3233/JAD-2011-110977. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
