

This is a preprint.
Graft incompatibility between pepper and tomato can be attributed to genetic incompatibility between diverged immune systems
- PMID: 38617251
- PMCID: PMC11014474
- DOI: 10.1101/2024.03.29.587379
Graft incompatibility between pepper and tomato can be attributed to genetic incompatibility between diverged immune systems
Update in
-
Graft incompatibility between pepper and tomato elicits an immune response and triggers localized cell death.Hortic Res. 2024 Sep 11;11(12):uhae255. doi: 10.1093/hr/uhae255. eCollection 2024 Dec. Hortic Res. 2024. PMID: 39664688 Free PMC article.
Abstract
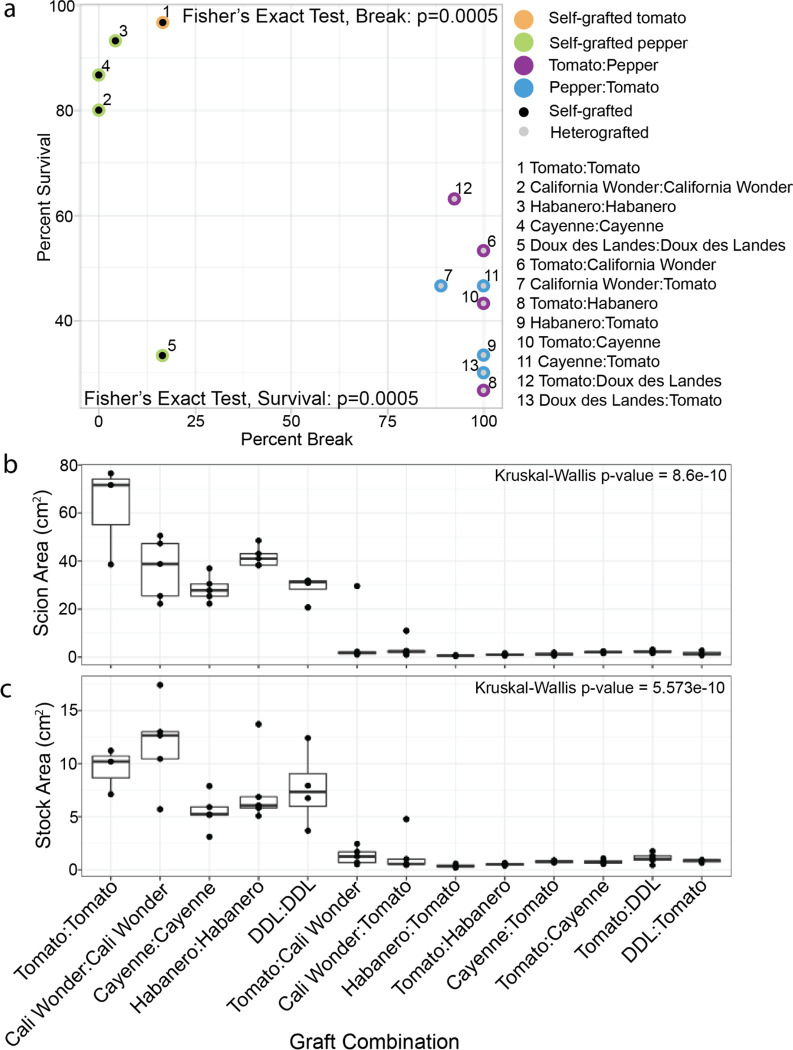
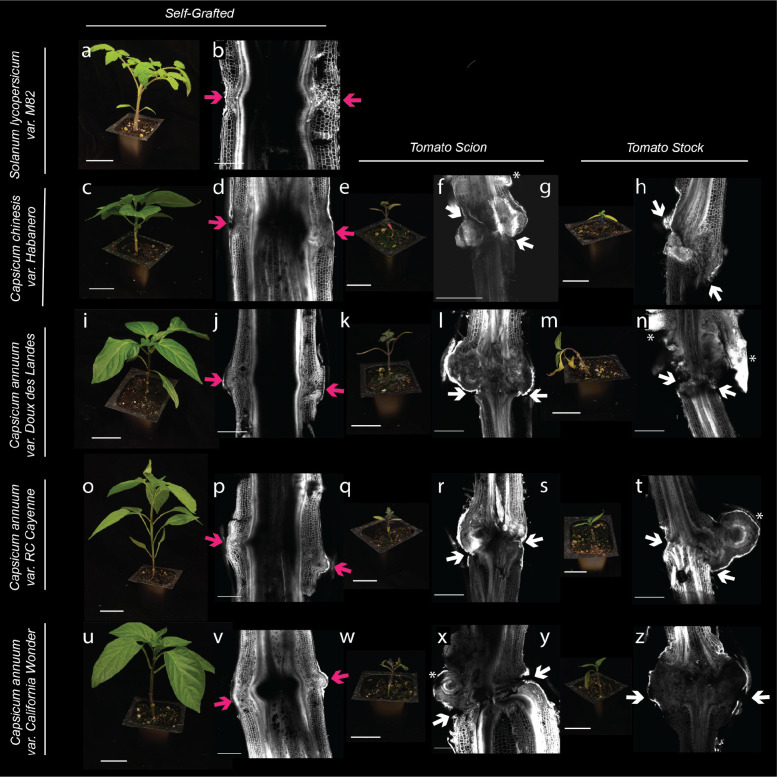
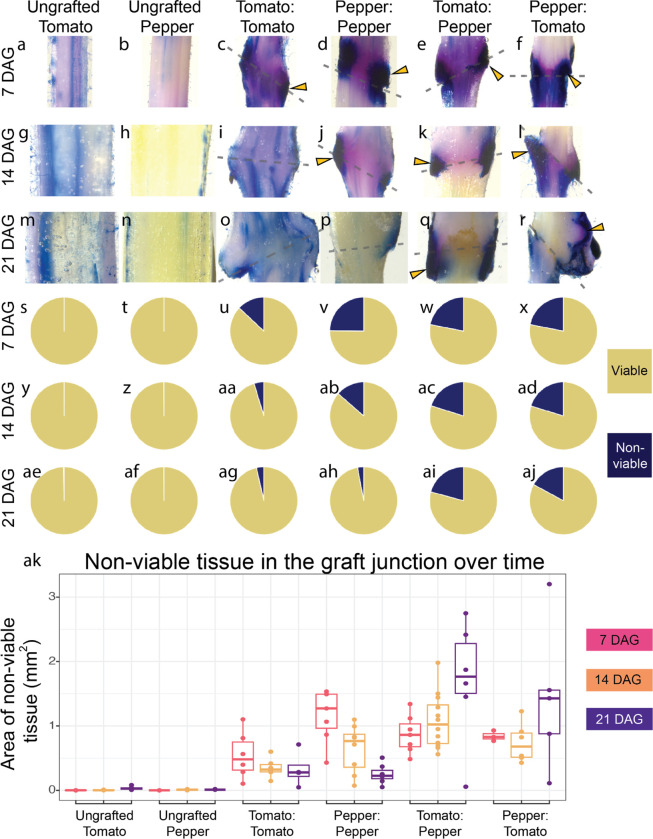
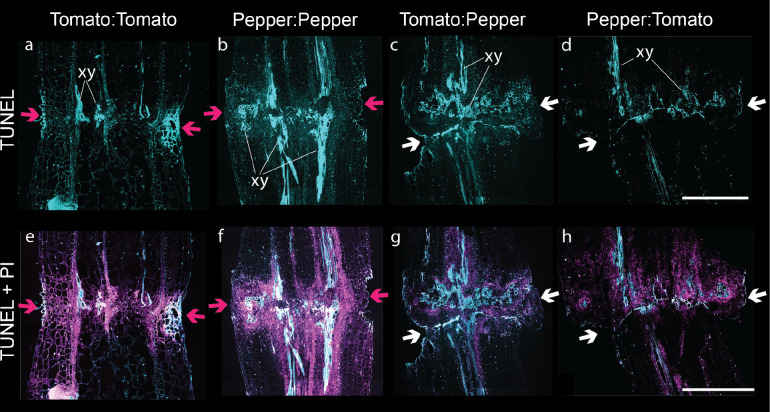
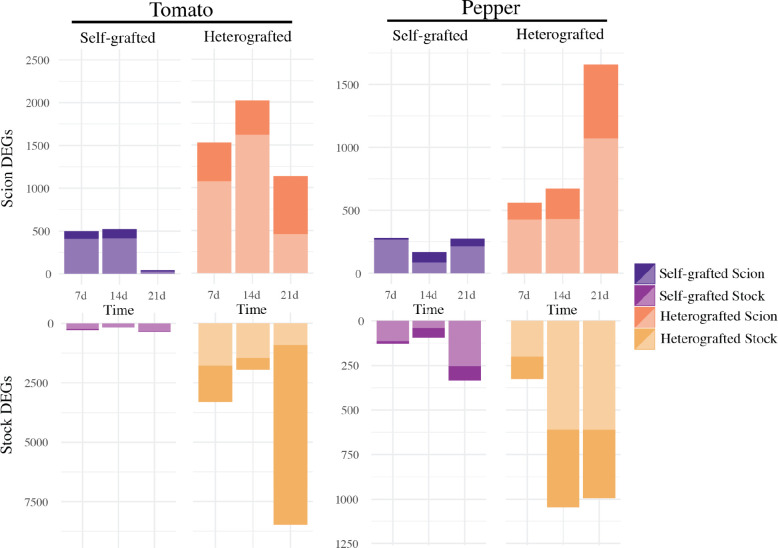
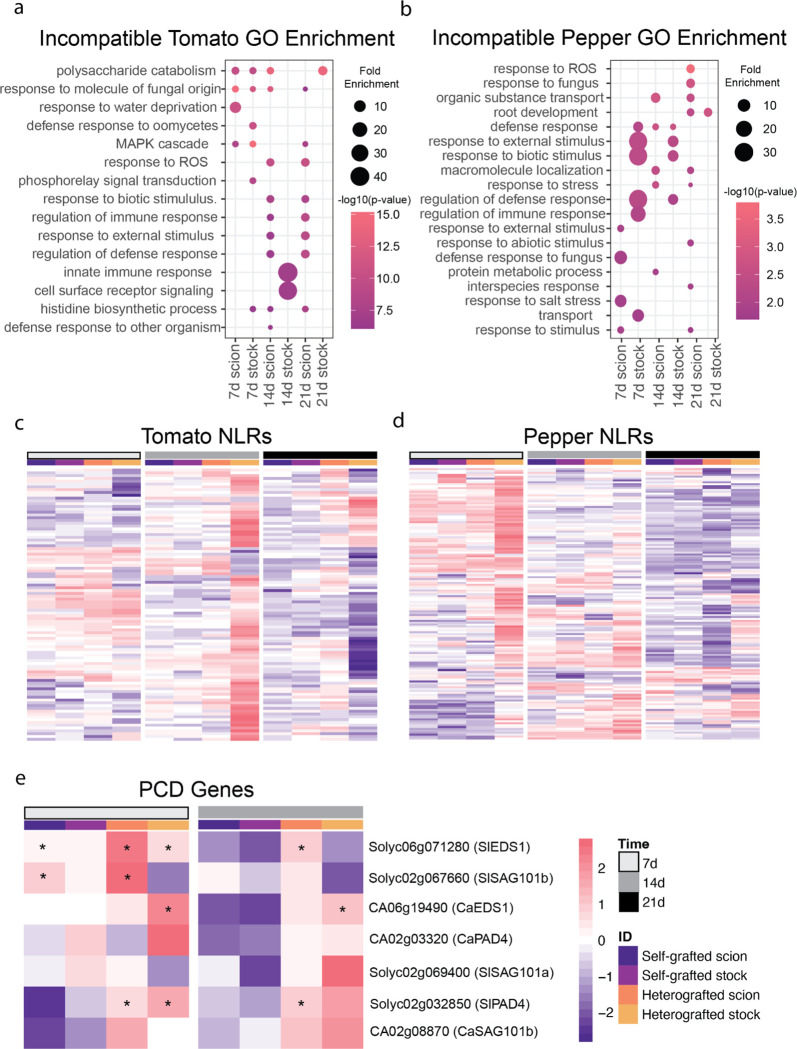
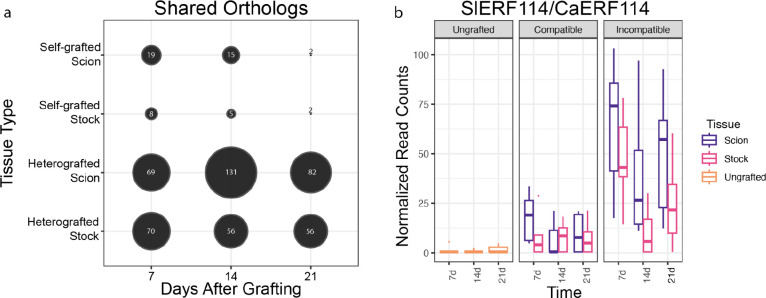
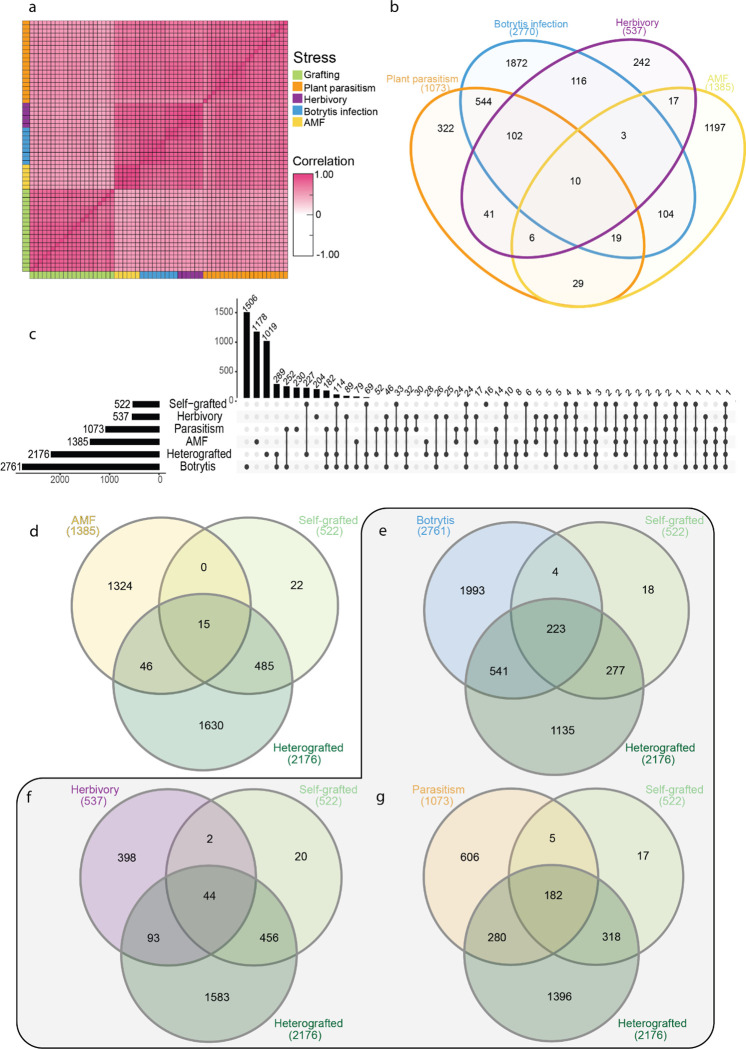
Graft compatibility is the capacity of two plants to form cohesive vascular connections. Tomato and pepper are incompatible graft partners; however, the underlying cause of graft rejection between these two species remains unknown.We diagnosed graft incompatibility between tomato and diverse pepper varieties based on weakened biophysical stability, decreased growth, and persistent cell death using trypan blue and TUNEL assays. Transcriptomic analysis of cell death in the junction was performed using RNA-sequencing, and molecular signatures for incompatible graft response were characterized based on meta-transcriptomic comparisons with other biotic processes.We show that tomato is broadly incompatible with diverse pepper cultivars. These incompatible graft partners activate prolonged transcriptional changes that are highly enriched for defense processes. Amongst these processes was broad NLR upregulation and hypersensitive response. Using transcriptomic datasets for a variety of biotic stress treatments, we identified a significant overlap in the genetic profile of incompatible grafting and plant parasitism. In addition, we found over 1000 genes that are uniquely upregulated in incompatible grafts.Based on NLR overactivity, DNA damage, and prolonged cell death we have determined that tomato and pepper graft incompatibility is likely caused by a form of genetic incompatibility, which triggers a hyperimmune-response.
Keywords: Solanaceae; autoimmunity; graft compatibility; hypersensitive response; plant grafting; programmed cell death.
Conflict of interest statement
Competing interests None to declare.
Figures
References
-
- Abdelkareem A, Thagun C, Nakayasu M, Mizutani M, Hashimoto T, Shoji T. 2017. Jasmonate-induced biosynthesis of steroidal glycoalkaloids depends on COI1 proteins in tomato. Biochemical and biophysical research communications 489: 206–210. - PubMed
-
- Andrews PK, Marquez CS. 2010. Graft Incompatibility. Horticultural Reviews: 183–232.
-
- Argles GK. 1937. A Review of the Literature on Stock-scion Incompatibility in Fruit Trees: With Particular Reference to Pome and Stone Fruits.
-
- Asahina M, Azuma K, Pitaksaringkarn W, Yamazaki T, Mitsuda N, Ohme-Takagi M, Yamaguchi S, Kamiya Y, Okada K, Nishimura T, et al. 2011. Spatially selective hormonal control of RAP2.6L and ANAC071 transcription factors involved in tissue reunion in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America 108: 16128–16132. - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources