

Trace gas oxidation sustains energy needs of a thermophilic archaeon at suboptimal temperatures
- PMID: 38622143
- PMCID: PMC11018855
- DOI: 10.1038/s41467-024-47324-2
Trace gas oxidation sustains energy needs of a thermophilic archaeon at suboptimal temperatures
Abstract
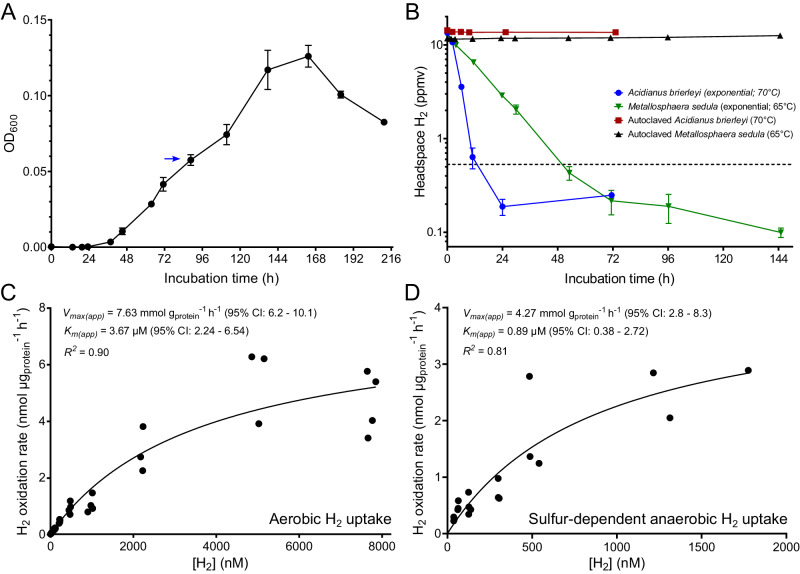
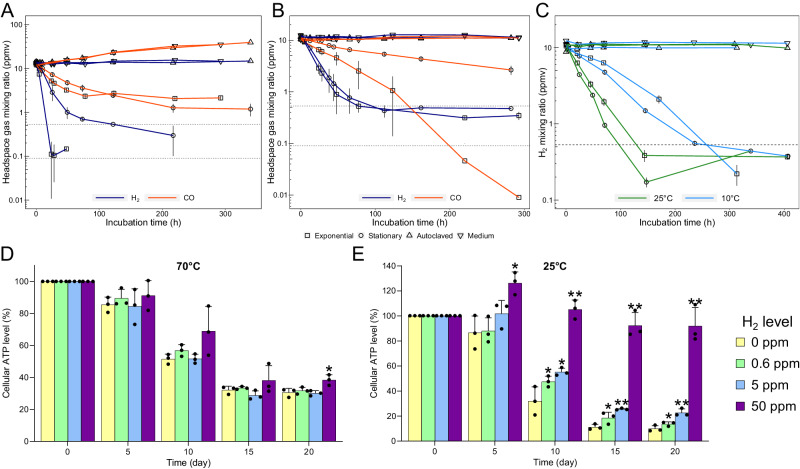
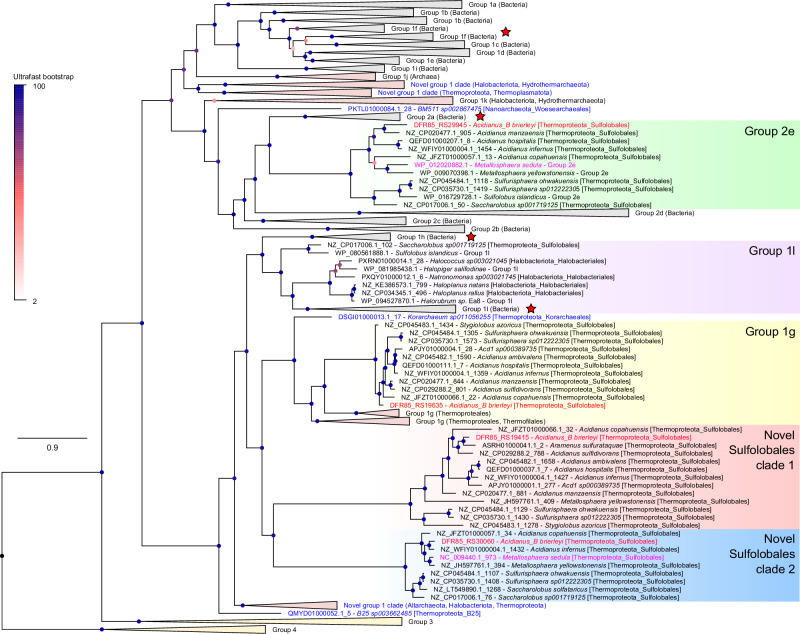
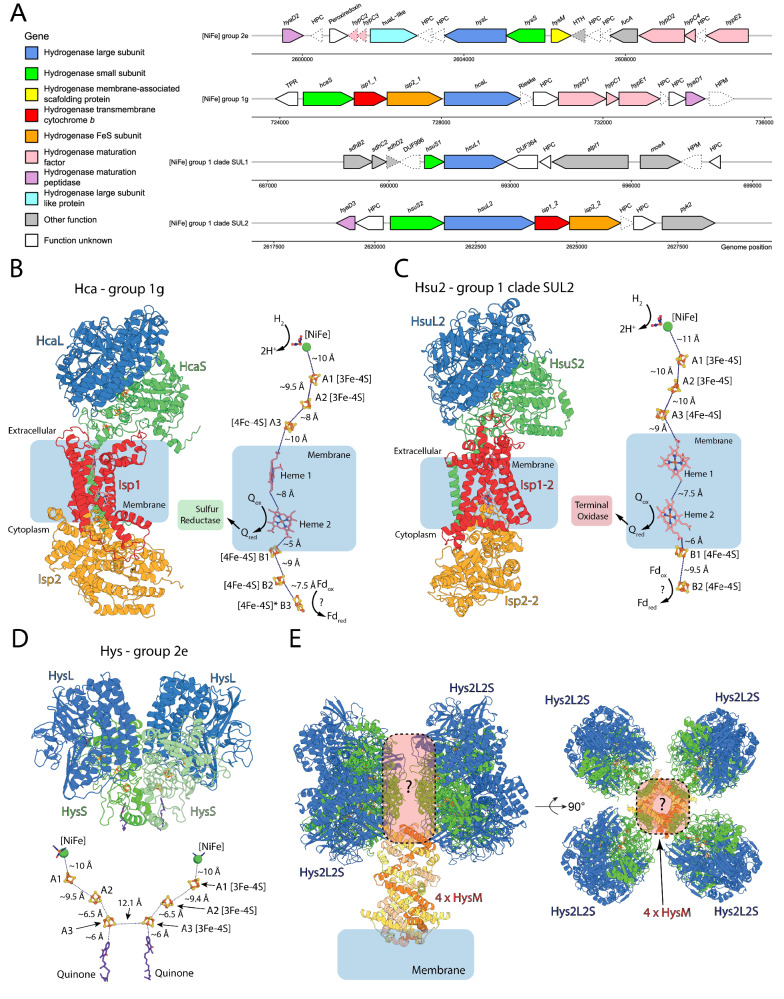
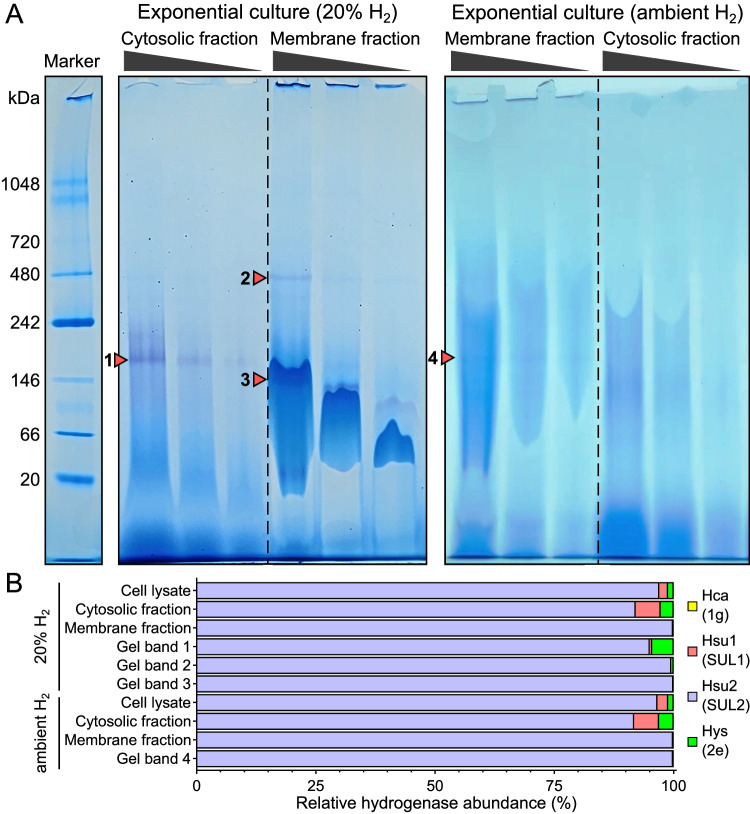
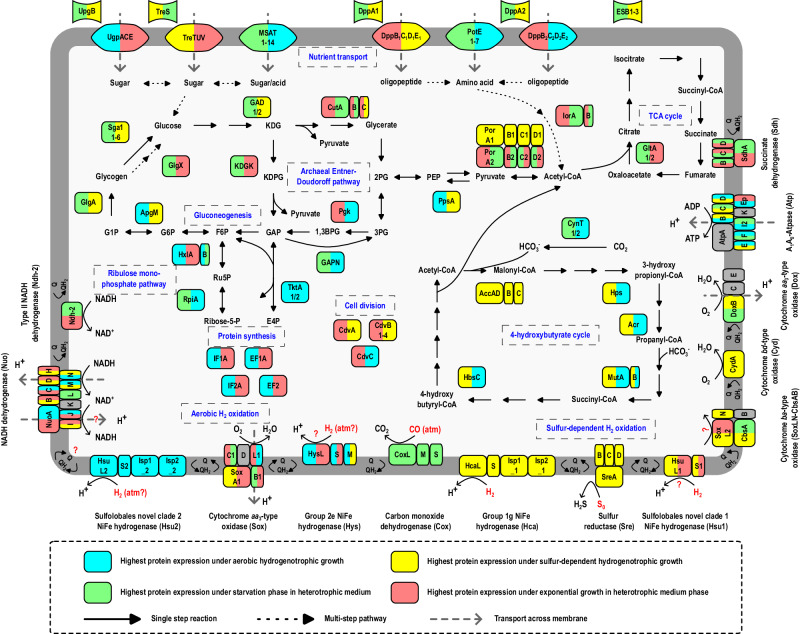
Diverse aerobic bacteria use atmospheric hydrogen (H2) and carbon monoxide (CO) as energy sources to support growth and survival. Such trace gas oxidation is recognised as a globally significant process that serves as the main sink in the biogeochemical H2 cycle and sustains microbial biodiversity in oligotrophic ecosystems. However, it is unclear whether archaea can also use atmospheric H2. Here we show that a thermoacidophilic archaeon, Acidianus brierleyi (Thermoproteota), constitutively consumes H2 and CO to sub-atmospheric levels. Oxidation occurs across a wide range of temperatures (10 to 70 °C) and enhances ATP production during starvation-induced persistence under temperate conditions. The genome of A. brierleyi encodes a canonical CO dehydrogenase and four distinct [NiFe]-hydrogenases, which are differentially produced in response to electron donor and acceptor availability. Another archaeon, Metallosphaera sedula, can also oxidize atmospheric H2. Our results suggest that trace gas oxidation is a common trait of Sulfolobales archaea and may play a role in their survival and niche expansion, including during dispersal through temperate environments.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Novelli PCC, et al. Molecular hydrogen in the troposphere: global distribution and budget. J. Geophys. Res. Atmos. 1999;104:30427–30444. doi: 10.1029/1999JD900788. - DOI
-
- Constant P, Chowdhury SP, Hesse L, Conrad R. Co-localization of atmospheric H2 oxidation activity and high affinity H2-oxidizing bacteria in non-axenic soil and sterile soil amended with Streptomyces sp. PCB7. Soil Biol. Biochem. 2011;43:1888–1893. doi: 10.1016/j.soilbio.2011.05.009. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
