

Characterization of Amycolatopsis 75iv2 dye-decolorizing peroxidase on O-glycosides
- PMID: 38625022
- PMCID: PMC11107159
- DOI: 10.1128/aem.00205-24
Characterization of Amycolatopsis 75iv2 dye-decolorizing peroxidase on O-glycosides
Abstract
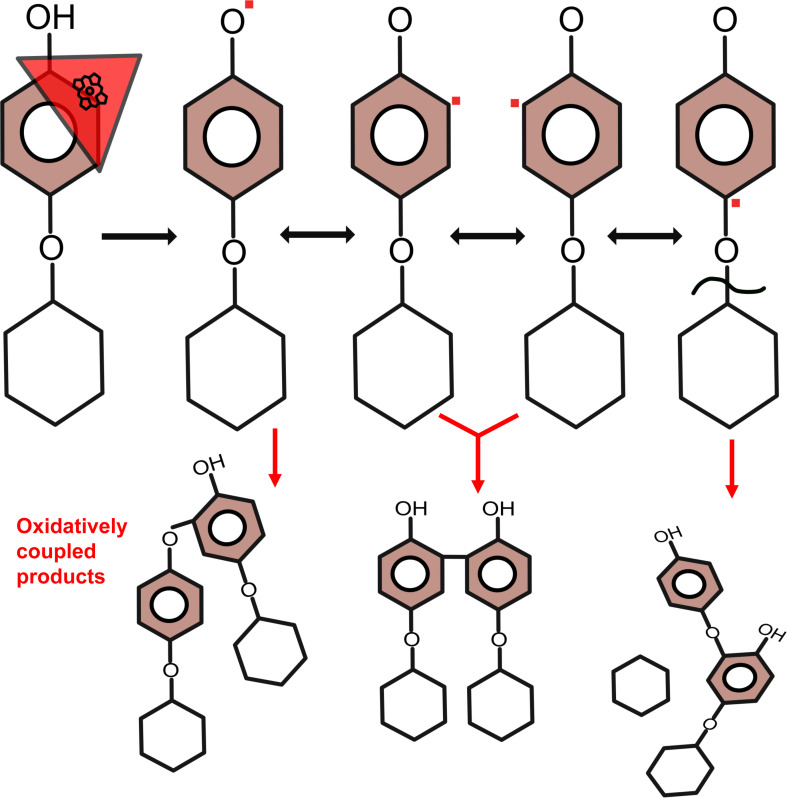
Dye-decolorizing peroxidases are heme peroxidases with a broad range of substrate specificity. Their physiological function is still largely unknown, but a role in the depolymerization of plant cell wall polymers has been widely proposed. Here, a new expression system for bacterial dye-decolorizing peroxidases as well as the activity with previously unexplored plant molecules are reported. The dye-decolorizing peroxidase from Amycolatopsis 75iv2 (DyP2) was heterologously produced in the Gram-positive bacterium Streptomyces lividans TK24 in both intracellular and extracellular forms without external heme supplementation. The enzyme was tested on a series of O-glycosides, which are plant secondary metabolites with a phenyl glycosidic linkage. O-glycosides are of great interest, both for studying the compounds themselves and as potential models for studying specific lignin-carbohydrate complexes. The primary DyP reaction products of salicin, arbutin, fraxin, naringin, rutin, and gossypin were oxidatively coupled oligomers. A cleavage of the glycone moiety upon radical polymerization was observed when using arbutin, fraxin, rutin, and gossypin as substrates. The amount of released glucose from arbutin and fraxin reached 23% and 3% of the total substrate, respectively. The proposed mechanism suggests a destabilization of the ether linkage due to the localization of the radical in the para position. In addition, DyP2 was tested on complex lignocellulosic materials such as wheat straw, spruce, willow, and purified water-soluble lignin fractions, but no remarkable changes in the carbohydrate profile were observed, despite obvious oxidative activity. The exact action of DyP2 on such lignin-carbohydrate complexes therefore remains elusive.
Importance: Peroxidases require correct incorporation of the heme cofactor for activity. Heterologous overproduction of peroxidases often results in an inactive enzyme due to insufficient heme synthesis by the host organism. Therefore, peroxidases are incubated with excess heme during or after purification to reconstitute activity. S. lividans as a production host can produce fully active peroxidases both intracellularly and extracellularly without the need for heme supplementation. This reduces the number of downstream processing steps and is beneficial for more sustainable production of industrially relevant enzymes. Moreover, this research has extended the scope of dye-decolorizing peroxidase applications by studying naturally relevant plant secondary metabolites and analyzing the formed products. A previously overlooked artifact of radical polymerization leading to the release of the glycosyl moiety was revealed, shedding light on the mechanism of DyP peroxidases. The key aspect is the continuous addition, rather than the more common approach of a single addition, of the cosubstrate, hydrogen peroxide. This continuous addition allows the peroxidase to complete a high number of turnovers without self-oxidation.
Keywords: O-glycosides; dye-decolorizing peroxidase; recombinant expression.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
