

Structure and dynamics of a pentameric KCTD5/CUL3/Gβγ E3 ubiquitin ligase complex
- PMID: 38625940
- PMCID: PMC11047111
- DOI: 10.1073/pnas.2315018121
Structure and dynamics of a pentameric KCTD5/CUL3/Gβγ E3 ubiquitin ligase complex
Abstract
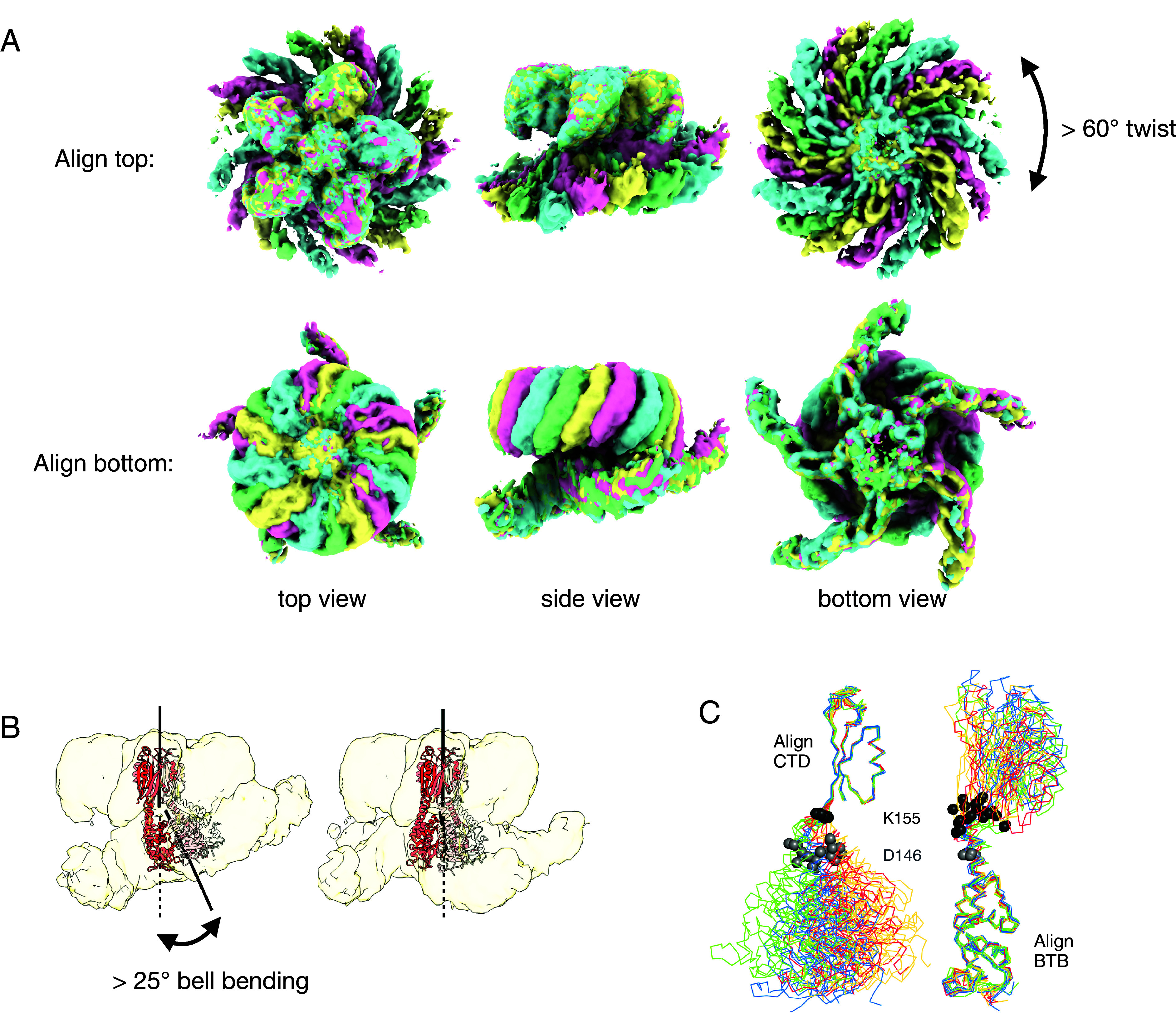
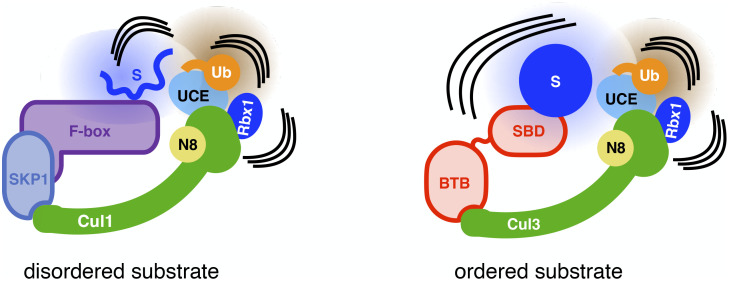
Heterotrimeric G proteins can be regulated by posttranslational modifications, including ubiquitylation. KCTD5, a pentameric substrate receptor protein consisting of an N-terminal BTB domain and a C-terminal domain, engages CUL3 to form the central scaffold of a cullin-RING E3 ligase complex (CRL3KCTD5) that ubiquitylates Gβγ and reduces Gβγ protein levels in cells. The cryo-EM structure of a 5:5:5 KCTD5/CUL3NTD/Gβ1γ2 assembly reveals a highly dynamic complex with rotations of over 60° between the KCTD5BTB/CUL3NTD and KCTD5CTD/Gβγ moieties of the structure. CRL3KCTD5 engages the E3 ligase ARIH1 to ubiquitylate Gβγ in an E3-E3 superassembly, and extension of the structure to include full-length CUL3 with RBX1 and an ARIH1~ubiquitin conjugate reveals that some conformational states position the ARIH1~ubiquitin thioester bond to within 10 Å of lysine-23 of Gβ and likely represent priming complexes. Most previously described CRL/substrate structures have consisted of monovalent complexes and have involved flexible peptide substrates. The structure of the KCTD5/CUL3NTD/Gβγ complex shows that the oligomerization of a substrate receptor can generate a polyvalent E3 ligase complex and that the internal dynamics of the substrate receptor can position a structured target for ubiquitylation in a CRL3 complex.
Keywords: BTB proteins; G proteins; cryo electron microscopy; ubiquitin ligase.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures




References
-
- Rusnac D.-V., Zheng N., Structural biology of CRL ubiquitin ligases. Adv. Exp. Med. Biol. 1217, 9–31 (2020). - PubMed
-
- Wang P., Song J., Ye D., CRL3s: The BTB-CUL3-RING E3 ubiquitin ligases. Adv. Exp. Med. Biol. 1217, 211–223 (2020). - PubMed
-
- Errington W. J., et al. , Adaptor protein self-assembly drives the control of a cullin-RING ubiquitin ligase. Structure 20, 1141–1153 (2012). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
